
INTRODUCTION
To date, various efforts have been made to identify biomarkers linked to fertility mechanisms as well as to predict and diagnose male infertility at the molecular level [1-3]. However, the efficiency of the biomarkers varies, and they are not validated enough to be applied in the animal industry and to humans. Most biomarkers have not been fully studied, to enable directly determining cell functions and identifying the underlying mechanisms. Recently, Ras-related proteins (Rab), which are members of the Ras superfamily of monomeric G proteins, were identified as validated biomarkers that could predict male fertility. In 2015, Kwon et al. reported the use of Rab2A for evaluation of boar fertility [1, 2]. Rab proteins are known to be involved in acrosomal biogenesis and exocytosis. Rab proteins play a role in preserving and establishing the Golgi structure, and the Golgi apparatus is derived from the acrosome during spermatogenesis [4-6]. In addition, Rab proteins are not only linked to acrosome formation and reaction but also to sperm motility capacitation conditions [5]. Thus, the importance of the Rab proteins for male reproduction has been emphasized by several studies to identify the function of Rab proteins in spermatozoa of humans and animals [5-8]. Moreover, recently, the mechanism of Rab protein signaling has been identified in spermatozoa [6].
Previous studies have shown that Rab proteins are associated with morphological and motion kinematic functions in spermatozoa [5,6]. Further, Rab2A is a validated biomarker that aids in the prognosis and diagnosis of boar fertility [1,2]. Therefore, this study was designed to prove the application of other Rab proteins in predicting boar fertility. In the present study, the expression of Rab proteins (Rab3A, 4, 5, 8A, 9, 14, 25, 27A, and 34A) in 31 boar spermatozoa was evaluated before and after capacitation. Statistical analysis was performed to assess the correlation between Rab protein expression and in vivo fertility. In addition, the sensitivity, specificity, negative predictive value, positive predictive value, and overall accuracy of the Rab proteins were analyzed based on receiver operating curves (ROCs). Finally, artificial insemination was performed after predicting litter size based on the expression of Rab proteins, and the average litter size was evaluated.
MATERIALS AND METHODS
All processes were performed in accordance with the guidelines and were approved by the Institutional Animal Care and Use Committee of Kyungpook National University.
To produce F1 pigs, artificial insemination was performed at a commercial pig farm (Gyeongsan Swine Gene, Gyeongsan, Korea). Breeding environments (light, ventilation, and temperature) were controlled to exclude seasonal effects. Duroc semen were gathered by the gloved hand technique once per week [9], then diluted with Beltsville Thawing Solution for AI (20 × 109 sperm cells/100 mL). In vivo fertility data of 123 Duroc sows (mean age: 29.5 ± 0.53; range: 20–39 months; farrowing rate = 91.37 ± 1.35) were provided by Gyeongsan Swine Gene. Since the litter size of first and older parities sows is generally lower than that of other parities [10], 2–5 parity Duroc sows were randomly selected and inseminated with semen collected from a boar. Trained technicians artificially inseminated sperm cells (20 × 109 sperm cells) twice per estrus in the cervix.
Semen samples were collected three times per Duroc boar in different periods from randomly selected 31 Duroc boars (average litter size = 13.76 ± 0.38). The semen samples were centrifuged at 500×g for 20 min with a Percoll (70% and 35% [v/v], Sigma, St Louis, MO, USA) [5,11,12]. Samples were incubated in medium 199 (containing sodium bicarbonate [2.2 g/L], D-glucose [3.05 mM], calcium lactate [2.92 mM], sodium pyruvate [0.91 mM], fetal bovine serum [10%], and heparin [10 µg/mL]; Sigma) for 60 min at 37°C under 5% CO2 in air, to induce capacitation [5].
The expression of Rab proteins in Duroc spermatozoa was determined by enzyme-linked immunosorbent assay (ELISA) [5, 11, 13]. Total proteins were extracted with rehydration buffer (urea [7 M], thiourea [2 M], 3-[(3-cholamidopropyl) dimethylammonio]-1-propane sulfonate [4%], octyl b-D-glucopyranoside [1%], PMSF [24 mM], dithiothreitol [1%], Triton X-100 [0.05 %], and bromophenol blue [0.002 %]; Sigma) at 4°C for 1 h as previously described [2,5,14]. The final concentration of total proteins was calculated using the Bradford protein-binding protocol [15]. Extracted protein (50 µg/well) was loaded into plates and incubated overnight at 4°C. The plates were washed with 0.05% Tween-20 (PBST) and blocked with a blocking solution (1% bovine serum albumin [BSA] in PBST) for 90 min at 37°C. The plates were incubated with anti-Rab3A, 4, 5, 8A, 9, 11, 14, 25, 27A, and 34 antibodies (1 : 5,000; Abcam, Cambridge, UK) for 90 min at 37°C. Then, the plates were incubated with goat anti-rabbit IgG H&L (HRP) antibody (1 : 5,000; Abcam) for 90 min at 37°C. Finally, tetramethylbenzidine solution (Sigma) was used to activate peroxidase for 15 min at room temperature (RT). Then, the activation was terminated with sulfuric acid (1 N). Rab protein signals (450 nm) were detected by a microplate reader (Gemini Em; Molecular Devices Corporation, Sunnyvale, CA, USA). We then excluded the background signal (0.038) from the detected all data.
Sensitivity, specificity, negative predictive value, and positive predictive value, have been used in screening tests to evaluate quality assessment [1,2,11,13]. Sensitivity was the percentage that was correctly identified as having litter size when predicted litter size ≥ 14 based on the expression level of Rab proteins. The specificity was the percentage that was correctly identified as having litter size when predicted litter size < 14 based on the expression level of Rab proteins. The negative predictive value was the percentage when more than the specific expression level of Rab proteins, litter size was actually predicted < 14 among results predicted litter size ≥ 14 or < 14. The positive predictive value was the percentage when equal to or less than the specific expression level of Rab proteins, litter size was actually predicted ≥ 14 among results predicted litter size ≥ 14 or < 14.
Statistical analyses were performed using SPSS (version 25.0, Chicago, IL, USA). Correlation between expression of Rab proteins and litter size were analyzed using Pearson correlation coefficients. The expression of Rab proteins as a function of litter size ≥ 14 or < 14 was evaluated using ROCs. The cut-off value calculated by ROCs was relation to maximized sensitivity and specificity [1,2,11,12]. Finally, Student’s two-tailed t-test was applied to compare litter size predicted by cut-off value. Differences were considered significant at p < 0.05. All data are expressed as the mean ± SEM.
RESULTS AND DISCUSSION
Rab proteins that are concerned in Golgi trafficking play important roles in preserving and establishing the Golgi structure in eukaryotic cells. [4-6, 16]. In particular, Rab proteins induce acrosome biogenesis from the Golgi apparatus during spermatogenesis [17-19], and they are involved in acrosome exocytosis after capacitation [7,20,21]. Recently, it was confirmed that Rab proteins are present in the sperm tail and head [6]. In addition, it has been reported that Rab protein mechanism in spermatozoa is independent of that in general eukaryotic cells [6]. Moreover, Rab proteins are directly correlated with motion parameters and capacitation status before and after capacitation [5]. Rab proteins play a critical role in male fertility by being involved in capacitation status and sperm motility. However, further validation is needed to evaluate the association between fertility and Rab proteins. Therefore, the present study aimed to discover biomarkers for the prediction and diagnosis of male fertility by an analysis of the correlation between the expression of Rab proteins and litter size following capacitation.
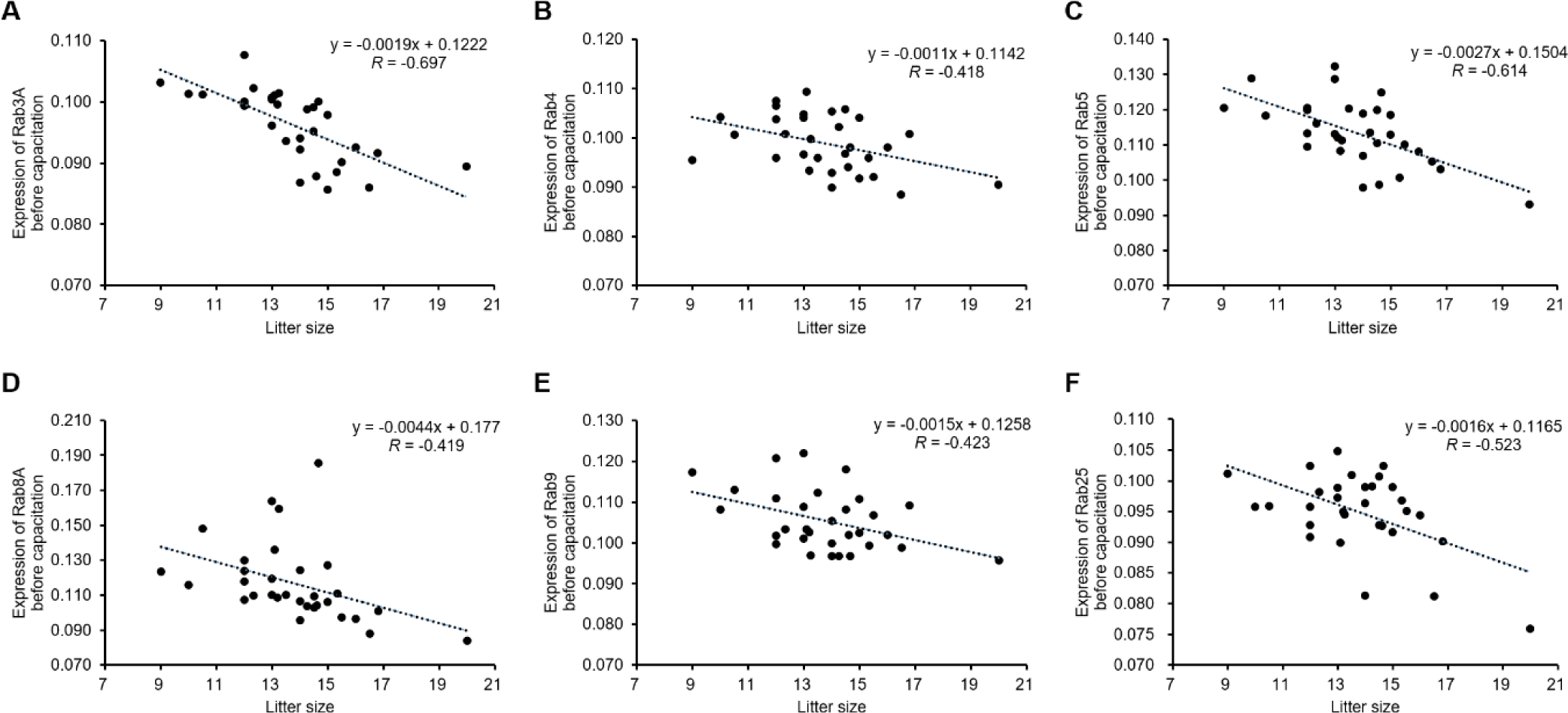
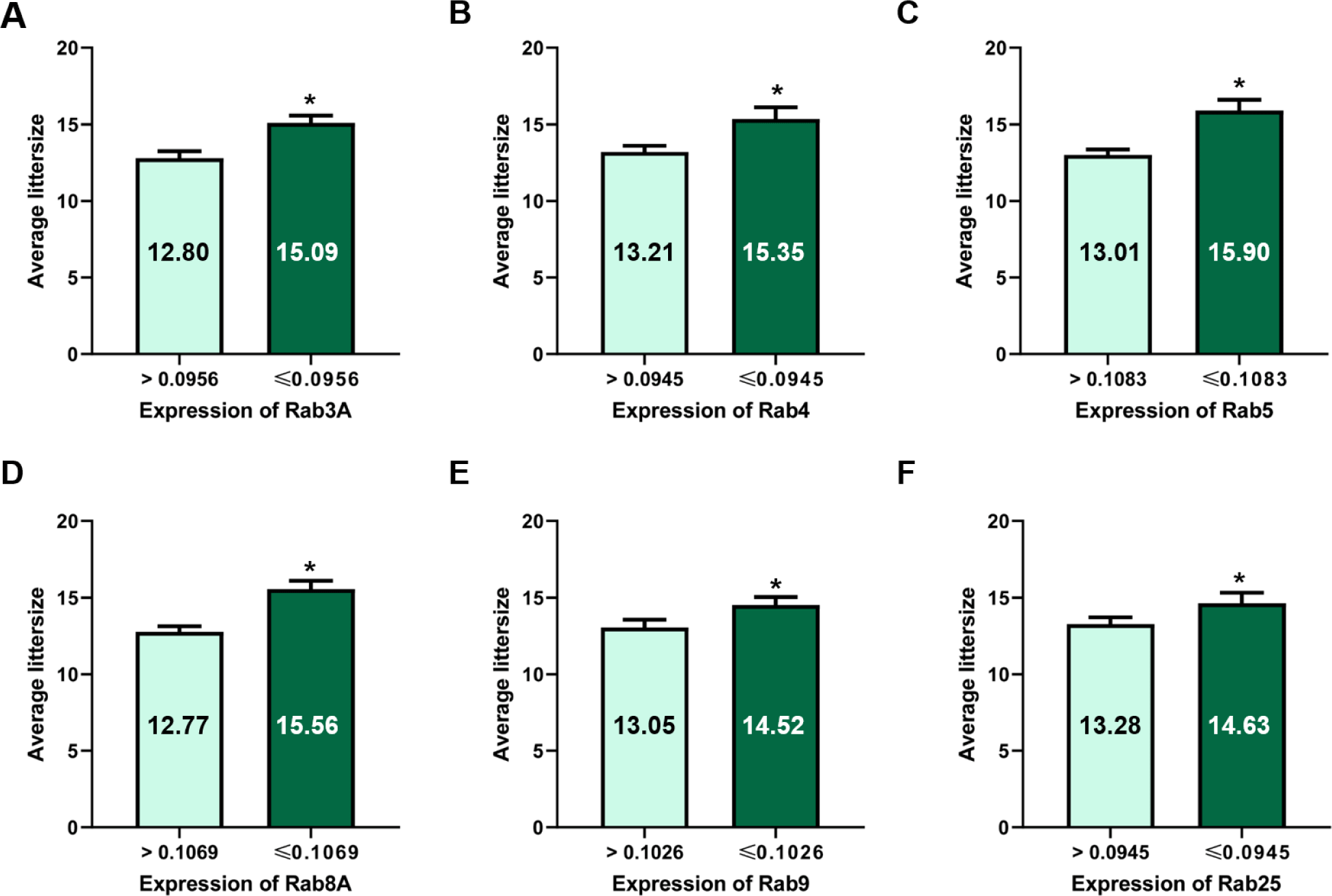
The expression of Rab3A (r = −0.697, p < 0.01), Rab4 (r = −0.418, p < 0.05), Rab5 (r = −0.614, p < 0.01), Rab8A (r = −0.419, p < 0.05), Rab9 (r = −0.423, p < 0.05), and Rab25 (r = −0.523, p < 0.01) was negatively correlated with litter size (Table 1 and Fig. 1). According to the ROCs, the cut-off values of Rab proteins for the 14 litter sizes were as follows: Rab3A = 0.0956, Rab4 = 0.0945, Rab5 = 0.1083; Rab8A = 0.1069, Rab9 = 0.1026; and Rab25, 0.0945 (Table 2 and Figs. 2 and 3). The average litter size with Rab3A > 0.0956 was 12.80 and ≤ 0.0956 was 15.09 (p < 0.05, Fig. 3A). The average litter size with Rab4 > 0.0945 was 13.21 and ≤ 0.0945 was 15.35 (p < 0.05, Fig. 3B). The average litter size with Rab5 > 0.1083 was 13.01 and ≤ 0.1083 was 15.90 (p < 0.05, Fig. 3C). The average litter size with Rab8A > 0.1069 was 12.77 and ≤ 0.1069 was 15.56 (p < 0.05, Fig. 3D). The average litter size with Rab9 > 0.1026 was 13.05 was ≤ 0.1026 was 14.52 (p < 0.05, Fig. 3E). The average litter size with Rab25 > 0.0945 was 13.28 was ≤ 0.0945 was 14.63 (p < 0.05, Fig. 3F). In addition, the overall accuracies of Rab3A, 4, 5, 8A, 9, and 25 before capacitation were 77.42, 67.74, 74.19, 83.87, 67.74, and 58.06 %, respectively (Table 2). The acrosome is derived from the Golgi apparatus and a lysosome-like structure is established during spermatogenesis [4, 16]. Because Rab proteins are involved in the establishment of the acrosome by regulating Golgi trafficking, they are key proteins in acrosome formation [4-6, 16]. Almost all Rab proteins are expressed in the sperm head and tail [6, 7, 22]. Further, Rab4 is correlated with the integrity of the sperm head structure, and Rab5 is correlated with various motion parameters in spermatozoa [5]. Our results showed that several Rab proteins (Rab3A, 4, 5, 8A, 9, and 25) are correlated with litter size before capacitation. Rab proteins play key roles in male fertility as they are involved in establishing sperm cell structure and function, including the structural integrity and motility of sperms. Moreover, five Rab proteins (Rab3A, 4, 5, 8A, 9, and 25) were validated based on their expression to predict fertility in the present study. It was confirmed that the litter size increased with the specific expression of Rab protein. Therefore, Rab3A, 4, 5, 8A, 9, and 25 may be used as new biomarkers for the prediction and diagnosis of male fertility. Particularly, Rab5 resulted in the highest increase in litter size and the third-highest overall accuracy. In addition, Rab8A resulted in the highest overall accuracy and the second-highest increase in litter size. Therefore, Rab5 and Rab8A are anticipated to be useful for developing efficient fertility-related biomarkers.
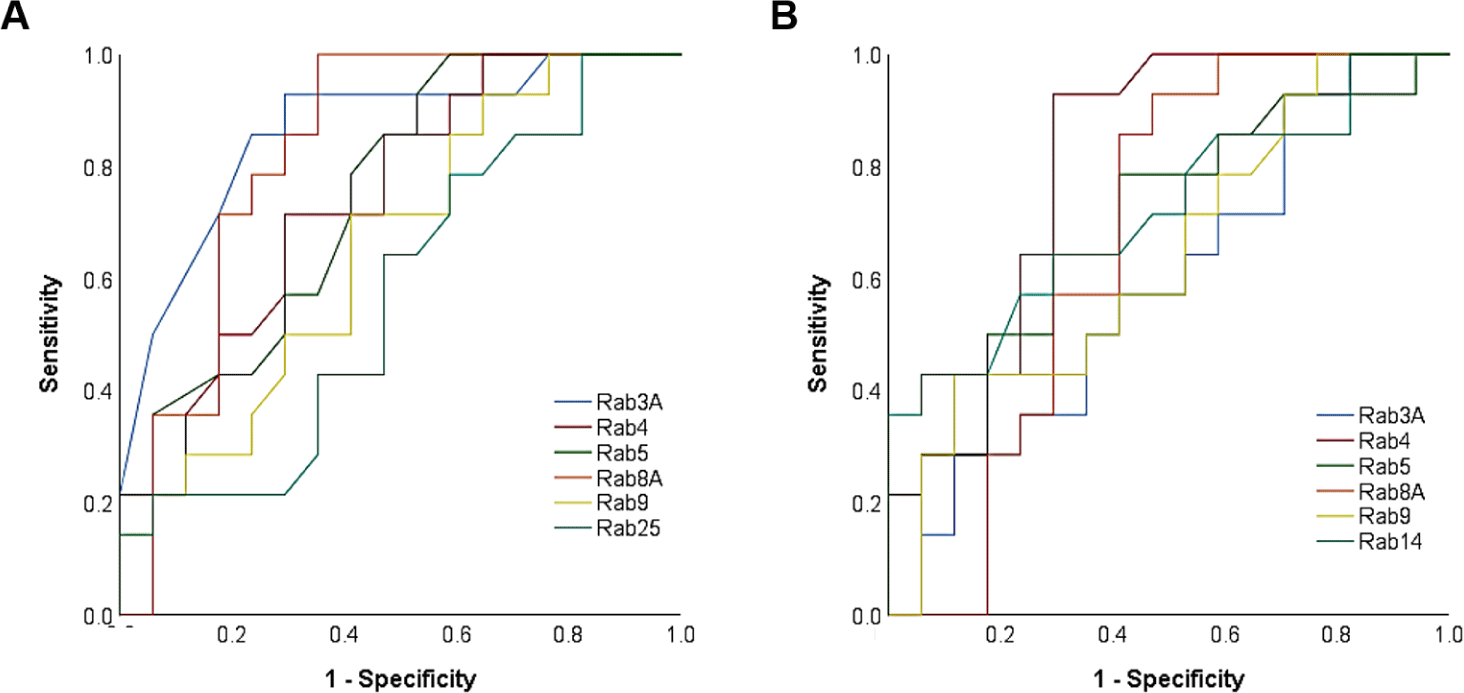
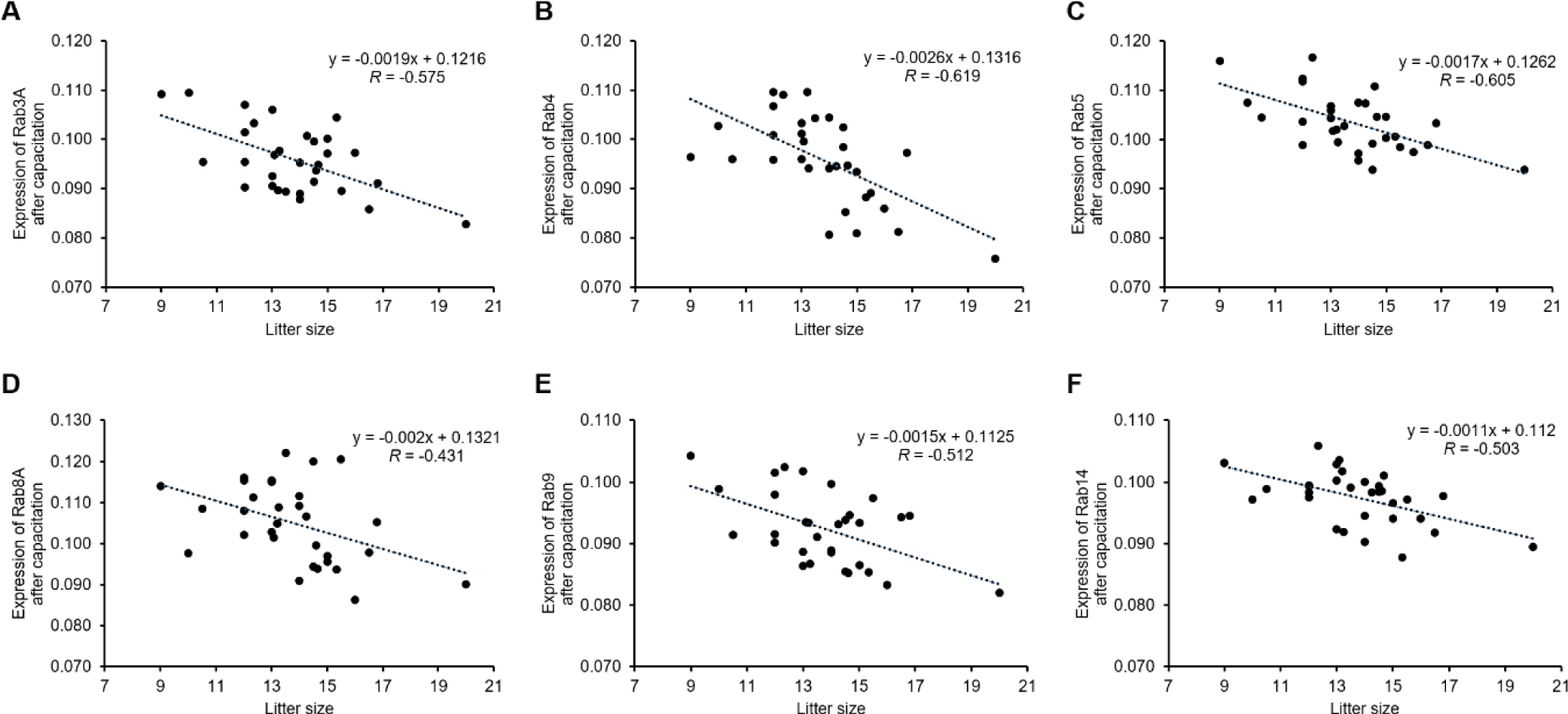
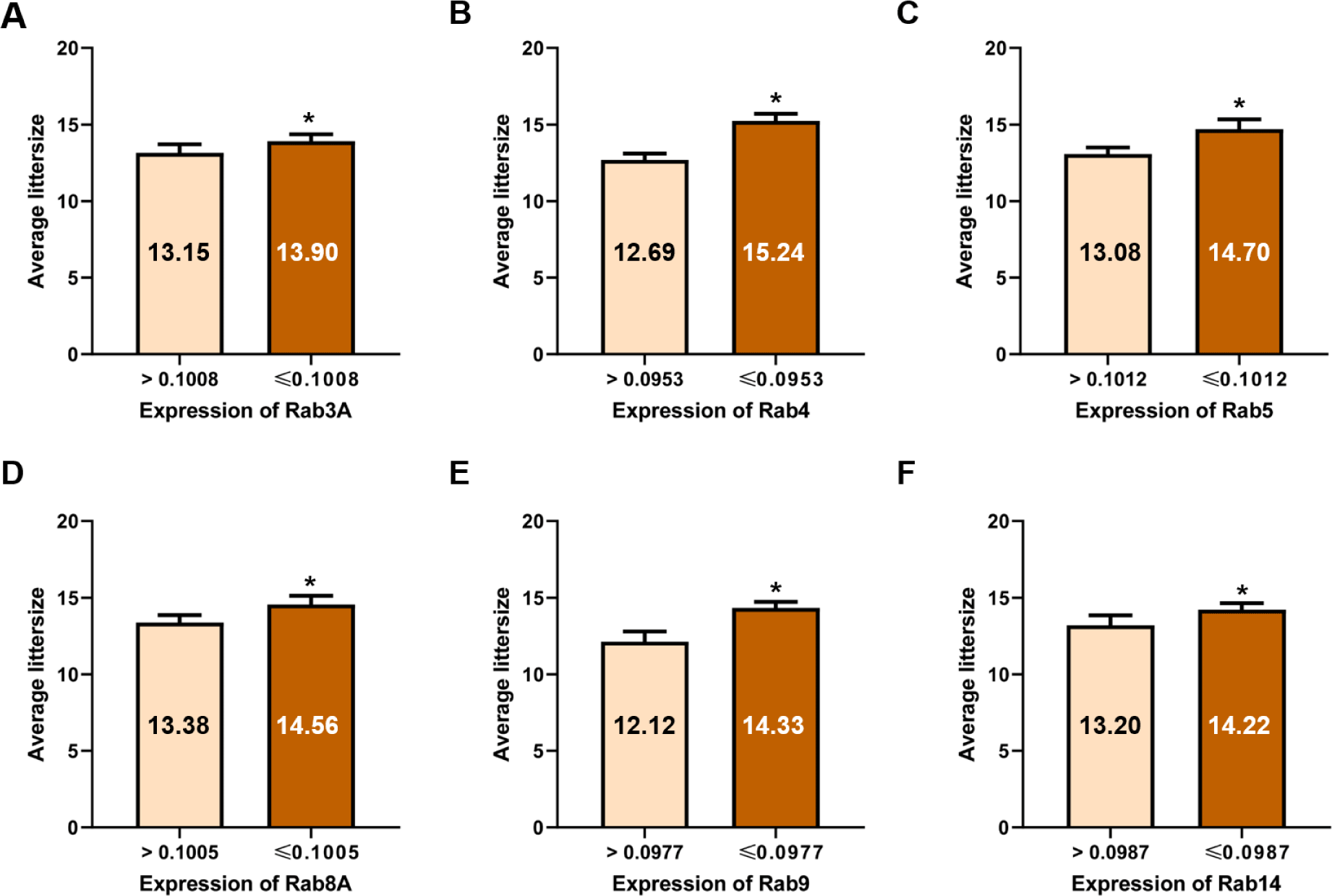
After inducing capacitation, most Rab proteins, except Rab25, were negatively correlated with litter size, similar to what was observed before capacitation. The expression of Rab3A (r = −0.575, p < 0.01), Rab4 (r = −0.619, p < 0.01), Rab5 (r = −0.605, p < 0.01), Rab8A (r = −0.431, p < 0.05), and Rab9 (r = −0.512, p < 0.01) was negatively correlated with litter size after capacitation (Table 3 and Fig. 4). Rab14 expression was negatively correlated after capacitation (r = −0.503, p < 0.01, Table 3 and Fig. 4). According to the ROCs, the cut-off values of Rab proteins for the 14 litter sizes were as follows: Rab3A = 0.1008, Rab4 = 0.0953, Rab5 = 0.1012, Rab8A = 0.1005, Rab9 = 0.0977, and Rab14, 0.0987 (Table 4 and Figs. 2 and 5). The average litter size with Rab3A > 0.1008 was 13.15 and ≤ 0.1008 was 13.90 (p < 0.05, Fig. 5A). The average litter size with Rab4 > 0.0953 was 12.69 and ≤ 0.0953 was 15.24 (p < 0.05, Fig. 5B). The average litter size with Rab5 > 0.1012 was 13.08 and ≤ 0.1012 was 14.70 (p < 0.05, Fig. 5C). The average litter size with Rab8A > 0.1005 was 13.38 and ≤ 0.1005 was 14.56 (p < 0.05, Fig. 5D). The average litter size with Rab9 > 0.0977 was 12.12 and ≤ 0.0977 was 14.33 (p < 0.05, Fig. 5E). The average litter size with Rab14 > 0.0987 was 13.20 and ≤ 0.0987 was 14.22 (p < 0.05, Fig. 5F). In addition, overall accuracies of Rab3A, 4, 5, 8A, 9, and 14 after capacitation were 59.38, 83.87, 70.97, 75.76, 64.52, and 70.97 %, respectively (Table 4). Similar to ejaculated spermatozoa, most Rab proteins also exist after capacitation [6]. In particular, Rab3A is correlated with capacitation status after capacitation [5]. Rab3A is well known for regulating acrosome exocytosis at end of capacitation [7, 22, 23]. In addition, Rab5 and 14 correlate with sperm motion parameters after capacitation [5]. Therefore, it may be considered that the Rab proteins are also playing an important role in male fertility by association with sperm motility and capacitation status. In the present study, the expression of Rab3A, 4, 5, 8A, 9, and 14 correlated with litter size after capacitation. Therefore, these five Rab proteins may be used to predict and diagnose male fertility. The most efficient Rab protein was Rab4, which resulted in the highest increase in litter size and overall accuracy after capacitation. Therefore, our results suggest that Rab4 may be a strong fertility-related biomarker after capacitation to evaluate male fertility.
CONCLUSION
Taken together, our results showed that the expression of six Rab proteins before (Rab3A, 4, 5, 8A, 9, and 25) and after capacitation (Rab3A, 4, 5, 8A, 9, and 14) was correlated with litter size. In addition, an increase in litter size was confirmed while evaluating the ability of the individual Rab proteins to predict litter size based on the cut-off value calculated by ROCs. The Rab8A protein had the highest overall accuracy before capacitation and Rab4 had the highest overall accuracy after capacitation (both 83.87%). Our results suggest that Rab proteins are correlated with litter size and may be applied as fertility-related biomarkers. We anticipate that it may be possible to improve productivity in pigs, as well as other domestic animals, using Rab proteins as biomarkers to analyze male fertility and select superior sires. Moreover, it suggests that Rab proteins may be applied to humans for the prediction and diagnosis of male fertility, as well as for identifying the cause of idiopathic male infertility/subfertility in patients.