
INTRODUCTION
Weaning is a stressful period for young pigs because several stressors including separation from mother sow, diet form change, and environmental changes [1]. In addition, weaned pigs must acclimatize to less digestible plant-based solid feed rather than easily digestible sow milk [2]. These multiple stress factors cause loss of appetite, which leads to insufficient energy intake for the maintenance of gastrointestinal health [3]. Additionally, the damage of intestinal epithelial layers because of weaning stress reduces trans-mucosal resistance [2]. Therefore, piglets in the weaning period are sensitive to infection by enteric pathogens, such as Escherichia coli, which leads to diarrhea [4,5].
To prevent post-weaning diarrhea and to improve growth performance and gut health of piglets, various antibiotics have been used as growth promoters [6,7]. However, because of potential safety issues, such as antimicrobial resistance or antibiotic residue, that may occur when antibiotics are overused, most countries have banned the use of antibiotics as growth promoters [8,9]. Therefore, a number of studies are being conducted to develop powerful alternatives to antibiotics for maintaining the health of weaned pigs [10].
Probiotics are defined by the FAO/WHO group as “live microorganisms which, when administered in adequate amounts, confer a good health benefit on the host” [11,12]. It has been well studied that probiotics could be alternatives to antibiotics because of their ability to reduce diarrhea and improve the growth performance of weaned pigs [13]. General strains of probiotic spices are mostly isolated from consumers of probiotics, including humans, chickens, or pigs. Commonly, Bacillus, Lactobacillus, Bifidobacterium, Enterococcus, and Streptococcus are used as probiotics [11]. Among them, Bacillus is an attractive bacterium for administration as a probiotic in swine feed because of its ability to form spores, which allows the bacterium to withstand high-temperature and the low-pH of gastric juices [14]. Additionally, spore-forming bacteria, such as Bacillus subtilis and Bacillus amyloliquefaciens, allow for long-term storage without losing viability [15]. According to previous studies, supplementation of Bacillus has been shown to improve growth performance by enhancing gut barrier integrity and relieving post-weaning diarrhea [14,16].
Although several previous studies have shown that Bacillus has beneficial properties for the host, the results of its effects on the immune system or intestinal microbiota of weaned pigs are still inconsistent. Therefore, this study was conducted to investigate the effect of Bacillus-based probiotics on growth performance, nutrient digestibility, intestinal morphology, immune response and intestinal microbiota of weaned pigs.
MATERIALS AND METHODS
The procedure of experiment was reviewed and approved by the Institutional Animal Care and Use Committee of Chungnam National University, Daejeon, Korea (approval #CNU-00910). This study was carried out at the facility of Animal Research Center of Chungnam National University. A total of healthy 40 weaned pigs ([Landrace × Yorkshire] × Duroc; 7.01 ± 0.86 kg; 28 d old) were assigned to 2 treatments in a randomized complete block design (block = body weight [BW] and sex) with 4 pigs per pen and 5 replicated pens per treatment. The dietary treatment was either a typical nursery diet based on corn and soybean meal (CON) or CON supplemented with 0.01% probiotics (PRO). Both dietary treatments were formulated to meet or exceed the requirements of the National Research Council [17] of weaned pigs and to have comparable metabolizable energy, crude protein (CP), and lysine levels (Table 1). Animal plasma, antibiotics, and zinc oxide were excluded from the piglet diets to avoid antimicrobial or physiological effects, and all feeds were provided in meal form. The dietary probiotics used in this study were a commercial product containing a mixture of Bacillus subtilis and Bacillus licheniformis. Pigs were housed in an environmentally controlled room and each pen was equipped with a feeder and water. Also, piglets were allowed ad libitum access to feed and water throughout the experiment.
The BW of pigs was recorded at d 0 of the experiment and at the last day of the experiment. The amount of feed added per pen was recorded, and feed residue were weighed from each pen on the last day of the experiment. The difference between the last BW and the initial BW was divided by the number of experimental days to determine the average daily gain (ADG). The feed intake per piglet was divided by the number of experimental days and was determined as the average daily feed intake (ADFI). To obtain the feed efficiency, the gain to feed ratio (G:F) was calculated by dividing ADG by ADFI for each pen. Diarrhea scores of all piglets during the experiment were recorded daily for 2 weeks from the first day after weaning. The diarrhea score for each pig was visually assessed daily on a scale of 1 to 5 (1 = dry feces, 2 = normal feces, 3 = slightly moist feces, 4 = semiliquid feces, mild diarrhea, and 5 = watery feces, severe diarrhea). The frequency of diarrhea (%) was calculated by counting the number of days the pigs had a diarrhea score of 4 or greater [18]. Whole blood samples were collected from one randomly selected pig in each pen using the jugular vein and stored in ethylenediaminetetraacetic acid tubes with anticoagulant or serum tubes (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ, USA) to measure total white blood cells (WBC) counts or inflammatory cytokines on d 7 and 14. Pigs were each fed a dietary treatment containing 0.2% indigestible chromium oxide during the last week of the experimental period to determine the apparent ileal and total tract digestibility of nutrients. For the collection of fresh fecal samples, rectal palpation was performed from one randomly selected pig in each pen for the last 3 days after the 4-day adjustment period. The collected stool samples and diet samples were stored at −20°C until analysis. Fecal samples for metagenome analysis were freshly collected from three randomly selected pigs in each treatment group on the last day of the experiment by rectal palpation with a sterile cotton swab, and the samples were placed in a sterile tube. The samples were taken to the laboratory on ice and stored at –80°C in a deep freezer until further analysis. On the last day of the experiment, one randomly selected piglet in each pen was anesthetized using 2 mL of suxamethonium chloride (Succicholine®, Ilsung Pharm, Seoul, Korea) by an intramuscular injection. Immediately after anesthesia, pigs were euthanized by CO2 gas [19]. Digesta samples of the pig gastrointestinal tract were collected from the terminal ileum before the ileocecal valve and were stored at −20°C until the time of analysis. For histological analysis, 3-cm sections were taken from the middle of the duodenum, jejunum, and ileum. The tissue samples were gently flushed with 0.05 M phosphate buffered saline (pH 7.2) and were then fixed in 10% neutral buffered formalin.
Before chemical analysis, frozen ileal digesta, fecal samples, and feed samples were thawed, dried at 60°C for 72 h and finely ground. Dry matter (DM, method 930.15) and nitrogen (method 999.03) of all samples were analyzed according to the procedures outlined in the Association of Official Analytical Chemists [20]. The gross energy (GE) was analyzed for each sample using a bomb calorimeter (Parr 1281 Bomb Calorimeter, Parr Instrument, Moline, IL, USA), and nitrogen was measured using a Kjeltec 2300 Analyzer (Foss Tecator AB, Hoeganaes, Sweden). Chromium levels in the samples were measured by UV spectrophotometer (Hitachi Z-5000, Hitachi High-Technologies, Tokyo, Japan). The apparent total tract digestibility (ATTD) and apparent ileal digestibility (AID) of the DM, CP, and GE were calculated for each sample as described by Stein et al. [21]. The fixed tissue samples taken from the small intestine were embedded in paraffin wax, sectioned to 5 μm using a microtome, and stained with hematoxylin and eosin. Stained sections were examined using a charge-coupled device camera-equipped microscope (TE2000, Nikon, Tokyo, Japan) with digital imaging software for measurements and counts (NIS Elements BR; Nikon). The 15 longest and straightest villi and their associated crypts were selected to evaluate morphological changes [22]. The morphometric measurements of intestinal tissue were carried out as follows: villus height; crypt depth; villus height to crypt depth ratio (VH:CD); and the number of goblet cells. Whole blood samples were analyzed by an automated hematology analyzer (scil Vet abc hematology analyzer; scil animal care company,Viernheim, Germany) calibrated for porcine blood for total WBC counts. The blood samples for serum were allowed to clot for 2 h at room temperature and then overnight at 4°C followed by centrifugation at 3,000×g for 15 min at 4°C. Serum concentrations of cytokines and cortisol were measured by using a porcine-specific tumor necrosis factor-α (TNF-α) or transforming growth factor-β1 (TGF-β1) ELISA kit (R&D Systems, Minneapolis, MN, USA), a porcine-specific cortisol ELISA kit (Cusabio, Wuhan, China), a porcine C-reactive protein or an immunoglobulin ELISA kit (Abnova, Taipei City, Taiwan), and measurements were conducted according to the manufacturer’s protocols. The optical density was measured at 450 nm, and all concentrations were calculated using a standard curve.
DNA was extracted from fecal samples using a PowerSoil DNA Isolation kit (MO BIO Laboratories, Carlsbad, CA, USA) according to the manufacturer’s protocol. DNA concentration and quality were determined by measuring absorbance at 230, 260, and 280 nm using a spectrophotometer (SpectraMax ABS Plus, Molecular Devices, San Jose, CA, USA). To amplify the V4 region from the 16S rRNA gene, 341F/805R primers (forward, 5′-CCT ACG GGN GGC WGC AG-3′; reverse, 5′-GAC TAC HVG GGT ATC TAA TCC-3′) were used. 16S rRNA gene amplicon sequencing was performed on the Illumina MiSeq platform at Macrogen (Seoul, Korea).
The raw paired-end reads were processed using Quantitative Insights Into Microbial Ecology 2 (QI- IME2; version 2020.6.0). Quality control (e.g., filtering, trimming the sequence and removing chimeric sequences) was performed on demultiplexed sequences with the DADA2 pipeline. Taxonomic assignment was performed with the Greengenes reference database (13_8 version) into operational taxonomic units (OTUs) with 97% similarity. To assess within-sample diversity, microbial α-diversity was calculated based on several metrics (Shannon, Chao1, and Simpson index) with the script “diversity α-group-significance” after making phylogenetic trees using the mafft and FastTree programs of QIIME2. To evaluate between-group diversity, β-diversity using phylogenetic-based weight and unweighted UniFrac distances were measured through the script “diversity beta-group-significance”. Additionally, based on the weight and unweighted UniFrac distance metrics, principal coordinate analysis (PCoA) plots were used to visualize β-diversity.
The PROC GLM procedures of SAS (SAS Inst., Cary, NC, USA) were used to analyze all data except frequency of diarrhea and fecal microbiota in a randomized complete block design, and the experimental unit was the pen. The statistical model for growth performance, nutrient digestibility, intestinal morphology, and immune responses of piglets included effects of dietary treatment as a fixed effect. The results are presented as the mean ± standard error of the mean. A chi-squared test was used for the diarrhea frequency. Statistical analysis of the fecal microbial composition was performed using QIIME2 and GraphPad Prism software version 7 (La Jolla, CA, USA). Permutational multivariate analysis of variance (PERMANOVA) was performed using QIIME2 to establish whether the profile of fecal microbiota between the two groups was significantly different and was based on UniFrac distance matrices. Statistical significance and tendency were considered at p < 0.05 and 0.05 ≤ p < 0.10, respectively.
RESULTS AND DISCUSSION
Weaned pigs fed PRO had significantly improved ADG (428.57 vs. 386.19 g/d; p < 0.05) (Table 2). There were no significant differences in ADFI, G:F, and frequency of diarrhea between CON and PRO. Additionally, as shown in Table 3, there were no differences in the AID of DM, CP, and GE and the ATTD of DM and GE between CON and PRO at the end of the experiment. Pigs fed PRO tended to have improved ATTD of CP (87.25% vs. 84.13%; p < 0.10). Several studies have shown that the addition of Bacillus to pig feed could improve growth performance or reduce the incidence of diarrhea [16,23]. However, some previous studies failed to observe such positive effects on growth performance [24–26]. These discrepancies in the effect of Bacillus on pig growth performance may be caused by various factors, such as diet formula, different Bacillus species, dose, pig age, or environmental features [15]. In this study, the AID of DM, CP, and GE and the ATTD of either DM or GE were not affected by Bacillus-based probiotics. However, pigs supplemented with Bacillus-based probiotics tended to show improved ATTD of CP. Based on these results, it can be speculated that improved growth rate of pigs may be a result of a tendency to increase ATTD in CP. Similarly, some previous studies demonstrated that pigs supplemented with Bacillus-based probiotics showed improved nutrient digestibility [27,28]. One of the major well-known benefits of Bacillusspp. is to eliminate pathogens through nutrient competition in the intestinal mucosa, thereby increasing the balance of host gut microbiota and maximizing nutrient utilization [29,30]. Additionally, Bacillus spp. can synthesize various types of extracellular enzymes and proteases [31,32]. Therefore, it is predicted that nutrient digestibility may be increased due to Bacillus-based probiotic supplements in feed.
Weaning leads to reduction of VH and VH:CD, impaired intestinal barrier function, and decreased mucin levels [2,3]. Intestinal epithelium cells play an important role in digestion, nutrient absorption, and protection from pathogens and toxins in the gastrointestinal tract [33,34]. Therefore, analyzing the morphology of the intestine is a profitable indicator for assessing the health and function of the intestine [35]. The goblet cells at the intestinal mucus layer produce secretory mucin glycoproteins or membrane-bound mucins, the proteins that form the mucus layer that acts as a line of defense against enteric pathogens and microbial adhesion and invasion [36,37]. In this study, the effects of Bacillus-based probiotics on the intestinal morphology of the small intestine (duodenum, jejunum, and ileum) and the number of goblet cells were evaluated (Table 4). Bacillus-based probiotics did not affect the intestinal morphology in the duodenum, jejunum, and ileum, but tended to increase the number of goblet cells in the ileum (16.91 vs. 10.83; p < 0.10). Similarly, several studies reported that small intestinal morphology was not affected by probiotic products [38–40]. However, other studies have demonstrated a positive effect on intestinal histo-morphology in pigs that consume Bacillus-based probiotics [37,41,42]. The inconsistent results may be because of different strains or levels of probiotics and the overall health of the pigs.
Probiotics are well known to regulate the production of lymphocyte cytokines and have a major effect on the immune system [43]. To investigate the effects of Bacillus-based probiotics on the immune systems of piglets, blood parameters were analyzed, including the number of WBC and serum cytokines. As shown in Table 5, the blood parameters were not significantly different between CON and PRO. The TGF-β1 in the serum tended to be higher in the probiotic group at d 7 (987.28 vs. 590.85 pg/mL; p < 0.10) and d 14 (1,095.56 vs. 636.03 pg/mL; p < 0.10). The TGF-β1 is an anti-inflammatory cytokine that is known to maintain intestinal epithelial integrity and support the differentiation of immature dendritic cells [44]. According to a previous study, administration of a probiotic mixture (Bacillus licheniformis and Bacillus subtilis) increased the expression of mRNA for the gene encoding TGF-β1 in ileum tissues of weaned pigs [45]. Multi-microbial probiotics also increased TGF-β concentration in the serum of sows [46]. The TNF-α is a pro-inflammatory cytokine that activates the secretion of other cytokines, macrophages, and neutrophils [47]. In addition, it plays an important role in the initial immune response by stimulating immature immune cells to become mature cells and enhancing the antigen-killing ability of these cells [48,49]. Previous studies have reported that feeding a probiotic mixture (Bacillus licheniformis and Bacillus subtilis) increased the mRNA expression of TNF-α, but did not show a significant change in the concentration of TNF-α in the serum of weaned pigs [45]. Similarly, in the current study, the concentration of TNF-α in the serum of the probiotic group tended to be higher on d 7 (366.82 vs. 317.94 pg/mL; p < 0.10), but the difference between the two groups disappeared on d 14 (386.42 vs. 341.43 pg/mL; p = 0.903). However, there was no significant difference between the two groups in the overall immunity index, which may be due to the minimal immune response of weaned pigs under good hygienic experimental conditions [28]. Consistent with this result, it was previously shown that the treatment with multi-strain probiotics had no effect on immune markers [50].
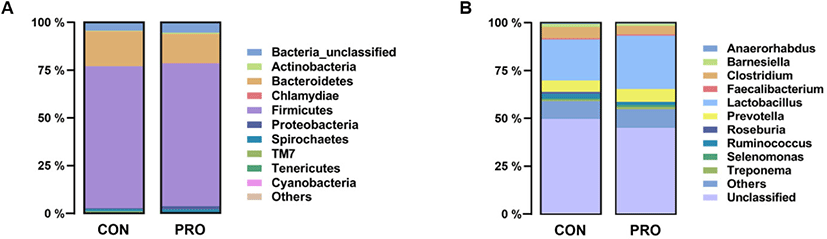
To investigate the effect of Bacillus-based probiotics on microbial composition, taxonomic analysis was performed. A total of 6 fecal samples yielded a total of 233,408 reads in this study, and the number of sequence reads ranged from 35,128 to 49,995 (mean = 38,901; median = 37,070). After quality control and removal of chimeras in OIIME2, the sequences were clustered into 1,799 OTUs based on 97% identity in our fecal samples. The relative abundances of the fecal microbiota of the control and probiotic groups at the phylum and genus levels are shown in Fig. 1. At the phylum level, the bacterial composition of the CON group on the last day of the experiment was predominated by the phyla Firmicutes (74.38%), Bacteroidetes (18.38%), and Spirochaetes (1.18%). Other phyla with relative abundances lower than 1% were Proteobacteria, Spirochaetes, Actinobacteria, and Tenericutes. By comparison, the core fecal microbiota at the phylum level of the probiotic group was Firmicutes (74.76%), Bacteroidetes (15.28%), Proteobacteria (1.63%), and Spirochaetes (1.5%). The low-abundance (< 1%) phyla were Actinobacteria, Chlamydiae, and Cyanobacteria. The abundances of unclassified bacteria from the fecal microbiota of the CON and PRO groups were 4.40% and 5.34%, respectively. A total of 107 genera were classified from all six samples at the genus level. The major genera of the CON group were Lactobacillus (21.50%), Prevotella (5.98%), Clostridium (5.91%), Ruminococcus (2.57%), Barnesiella (1.59%), and Roseburia (1.04%). Furthermore, the predominant taxa at the genus level of the PRO group were Lactobacillus (27.77%), Prevotella (6.71%), Clostridium (4.51%), Ruminococcus (1.71%), Barnesiella (1.13%), Treponema (1.17%), and Selenomonas (1.04%). The abundance of unclassified genera in each group was 49.26% and 44.75%, respectively. Intestinal microbes have various functions, such as the production of volatile fatty acids and vitamin K, digestion of cellulose, and regulation of the immune system. Because these functions are closely related to the gut health of pigs, they have been a major issue in recent years. [1,8]. Our findings show that, as in previous studies, Firmicutes and Bacteroidetes in fecal samples accounted for more than 90% of the phyla in both the CON and PRO groups [51,52]. Most unclassified genera belong to the Ruminococcaceae family, which is known to play a beneficial role in gut health by breaking down various polysaccharides and fibers and producing short-chain fatty acids [53,54]. The most dominant genera at the genus level in both the CON and PRO groups were Prevotella and Lactobacillus. Since weaned pigs generally consume grain-based carbohydrate-rich feed, it is presumed that Prevotella and Lactobacillus, which are involved in the fermentation of non-starch polysaccharides and the production of short-chain fatty acids, are dominated [55,56].
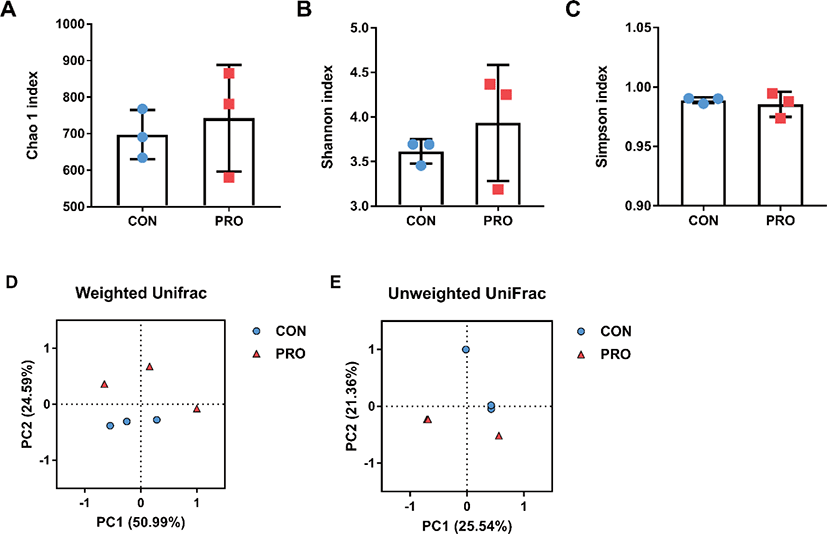
To determine microbial richness and evenness, Chao1, Shannon, and Simpson indices were calculated. The alpha diversity did not differ between the control and probiotic groups (Figs. 2A, B, and C, respectively). To compare the microbial diversity of fecal samples between the two groups, beta diversity was analyzed. PERMANOVA of weighted UniFrac distances showed that there was no remarkable difference between the two groups in microbial diversity (Fig. 2D). The PCoA based on unweighted UniFrac distances also indicated that there were no significant differences between the CON and PRO groups (Fig. 2E). A previous study showed that when Bacillus subtilis was fed to weaned pigs, there was no change in the alpha diversity of the small intestinal microbiota, but the microbial composition of the ileum and jejunum contents differed between the control and probiotic groups [57]. In addition, supplementing the feed with B. subtilis and/or yeast cultures did not affect the richness or diversity of feces from weaned pigs compared to the control group [58]. It was also reported that feeding weaned pigs higher doses of B. subtilis did not change the alpha diversity compared to the control group but caused a distinct separation between groups [41]. Additional studies are needed for various doses and strains of Bacillus-based probiotics that cause clear changes in the intestinal microbiota between groups.
CONCLUSION
The addition of Bacillus-based probiotics has beneficial effects on the growth performance including ADG and modulates immune system and gut microbiota of weaned pigs, suggesting that Bacillus could be utilized as a functional probiotic in animal feed. However, further study is needed to assess the effects of different Bacillus species and doses on the growth performance and intestinal health of weaned pigs and to evaluate the mechanism of action.