
INTRODUCTION
The poultry sector struggled after the ban on antibiotics to achieve improved feed utilization, minimized rate of mortality, and growth of healthy livestock [1]. To enhance the health and quality of eggs produced by the poultry sector, reliable feed additives should be introduced to the rations [2–4]. Supplements used to improve the production or animal health should have negligible side effects. Different herbal remedies might be used as potential substitutes for antibiotics and growth promoters. They are beneficial for health and have naturally occurring growth-promoting and anti-oxidative properties [5,6]. Plants contain a wide variety of active ingredients. Essential oils extracted from aromatic and medicinal herbs have anti-inflammatory, antibacterial, and digestive-stimulating characteristics, making them popular as poultry feed and other livestock supplements [7]. The ginseng plant and its extracts are natural growth stimulants that can replace antibiotics and have been used in Asian countries, such as Korea, Japan, and China, for over two millennia, leading to its global expansion in recent years [7]. Ginseng and its extracts have anti-allergy, anti-inflammatory, anti-cancer, anti-fatigue, anti-stress, and immunomodulatory properties in their essential oils, ginsenoside, polyacetylenic alcohol, saponin, peptides, polysaccharides, and vitamin contents, present in proportions based on the culture method used for growing the ginseng [8,9]. Based on the vegetative methods, ginseng can be classified as either wild or cultivated. Artificially grown ginseng is mostly collected after 5–6 years of methodical farming in an open field, while wild ginseng (WG) is sown in the deep mountain at a height between 800–1,500 meters. WG grows more slowly, is cheaper to maintain, and is more susceptible to environmental changes, favoring daily temperatures and less exposure to sunlight when compared with cultured ginseng. WG root has more medicinal effects than cultivated ginseng, which could be the major factor leading to differences in both types of ginsengs [10]. However, the roots from ginseng have not been widely and regularly used due to its higher cost and challenging cultivation. Nonetheless, cells from the plant have been used commercially in various foods and cosmetics due to the growth of the contemporary industry [11]. Artificial vegetation approaches have served as a solution to the mass production of ginseng adventitious roots with the same chemical content ratios as the native roots [12]. The main goal of our study was to evaluate the effect of Korean WG adventitious root derived from artificial vegetation on the laying performance, egg quality, ginsenoside concentration of yolk, cytokine, and microflora of laying hens.
MATERIALS AND METHODS
The experiment was carried out at the laying hen breeding facility of Kangwon National University with the approval of the University’s Animal Experimental Ethics Committee (KW-220413-1). In order to evaluate the effect of WG powder in layers diet, 90 pieces of 24 weeks old laying hens (Institut de selection Animale [ISA] Brown weighting 1,894 ± 0.12 g) were tested in a completely randomized design with 3 treatments and 5 repetition with 6 numbers per replicate. The birds were initially subjected to an adaptation period of 14 days, and the feeding experiment was conducted for a total of 12 weeks (phase 1, 0–4 weeks; phase 2, 5–8 weeks; phase 3, 9–12 weeks). The experiments were divided into the CON (basic feed), WG1 (basic feed + 0.1% WG), and WG2 (basic feed + 0.5% WG). The WG root was dried and roughly grinded into a powder form to release the active ingredients in the root using the CE commercial mini SUS304 electric 12–120 herb grinder model number LFJ-15B, and the inclusion level was based on economic reasons which is an important factor considered in our study. Previous researchers evaluated the effect of ginsengs between 1.0%–5.0% [5,13]. However, we tried to evaluate its potency at a lower dosage, but with WG plant grown for a period of 10 years. WG is generally believed to be more effective based on factors such as duration, specie, and their parts such as the leaves, root, or rhizomes supplemented in diets [13]. The lighting period was adjusted to 16 hours per day, and the chemical composition and mixing ratio of the experimental diet fed in this study were shown in Table 1, and the nutrient levels were formulated to meet or exceed the nutrient requirement as prescribed by ISA brown commercial management guide [14]. The feeds and water were administered at ad libitum in mash form, and all hens were housed in a window-less and environmentally controlled room, with room temperature kept between 20°C–22°C, and each cage was equipped with individual nipples, feeders, and nest enrichments according to the EU laying down minimum standards for the protection of laying hen [15].
Content in kg: vitamin A, 10,000 IU; vitamin D, 2,000 IU; vitamin E, 0.25 IU; vitamin K3, 2 mg; Vitamin B12, 10 mg; choline, 250 mg; folic acid, 1 mg; niacin, 30 mg; pantothenic acid, 10 mg; pyridoxine, 3 mg; riboflavin, 6 mg; thiamine, 2 mg; ethoxyquin, 125 mg; cobalt, 0.3 mg; copper, 10 mg; iron, 60 mg; iodine, 0.5 mg; manganese, 40 mg; selenium, 0.2 mg; zinc, 50 mg.
The layer hens were weighed after adaptation, and at the end of each phase (4, 8, and 12 weeks). Feed conversion ratio ([FCR] kg of feed/kg of eggs,) for all periods was calculated on a cage basis from egg production (hen day egg production, HDEP), average egg weight (AEW), and average daily feed intake (ADFI). Eggs were collected daily and egg production was expressed on a hen-day basis (% hens day). Individual egg weights were recorded and then used to calculate the mean egg weight for all experimental periods. The egg mass (EM) was calculated by multiplying egg weights and egg production rate [16].
Egg and eggshell quality examinations (Haugh units, yolk colour, yolk and albumin weights, yolk and albumin percentages, eggshell thickness, and hardness) were undertaken at the end of each phase. For measuring these parameters, eggs were collected on the last day of the phases. Haugh units, yolk colour, yolk, and albumin weights were measured by Egg multi-tester (Touhoku rhythm, Tokyo, Japan). Eggshell breaking strength was evaluated using a model II egg shell force gauge (Robotmation, Tokyo, Japan). A dial pipe gauge (Ozaki MFG, Tokyo, Japan) was employed for measuring the egg shell thickness, which was determined on the basis of the average thickness of the rounded end, pointed end, and the middle of the egg, excluding the inner membrane [16].
Five birds per treatment were selected for this process. A minimum of 5 mL blood was collected from the median underwing coverts and was withdrawn slowly using a disposable syringe and a needle (21 gauge) and transferred into a heparin tube on ice.
To isolate a PBMC, a ficoll solution was dispensed into the tube containing the blood then centrifuged for 40 min at 4000 rpm at 20°C using the Union 55R Refrigerated multipurpose centrifuge, Hanil Science Industrial. with the break set at 0. After separation, the final solution was stored at −4°C.
The RNeasy Mini kit (Qiagen, Hilden, Germany), was used for messenger ribonucleic acid (mRNA) extraction from the PBMC. In detailed, prior to homogenization using an Ultraturrax homogenizer (Polytron PT 1600E®, Kinematica, Luzern, Switzerland), the cells were trapped with 600 µL lysis-buffer (containing β-mercaptoethanol and RLT-buffer).
250 ng of extracted mRNA and 100 l of TaqMan Reverse Transcription Reagents® (Life Technologies, Carlsbad, CA, USA) were used to reverse transcribe the mRNA. 5 micro liter of random hexamers, 2 micro liter oligo-DT, 10 micro liter 10× bufer, 25 mM MgCl2, 20 µL dNTP, 2 µL RNAse inhibitor, and 6.2 µL of MultiScribe Reverse Transcriptase were employed as the reagents. Reverse transcription was carried out in an Eppendorf flexid, nexus gradient master cycler, SN:6332kl132036, Germany with annealing (25°C, 10 min), enzyme inactivation (95°C, 5 min). Relative tumor necrosis factor alpha (TNF-a) gene was quantified using 5 µL cDNA, 1.75 µL Aqua dest., 2 µL of TaqMan Master Mix and 0.25 µL of both forward and reverse primers, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was employed as the house keeping gene (Bioneer, Daejeon, Korea). The primer information is shown in Table 2. The qPCR Rotor-Gene Qiagen with Serial Number 0312272 (Corbett Research) with cycling condition 95°C followed by cycles of melting at 95°C for 15 s; annealing for the specified times and temperatures according to the primers; and extension at 72°C for a specified primer time and 40 cycles [17].
To measure fecal microflora differences according to each treatment, five fresh fecal samples per treatment were collected via the cloaca of layer hen at the end of each phase and immediately kept at −80°C until analysis which was carried out on the same day. The cloaca was gently palpated to stimulate the discharge of fresh fecal samples void of external contaminations from the pen into airtight containers. Deoxyribonucleic acid (DNA) extraction protocol according to QIAamp fast DNA stool mini kit Germany, cat. No. 51604 2016 was carried out. In Detail (Step 1), 200 mg stool was weighted with a scalpel into a 2 mL microcentrifuge tube and placed on ice. 1 mL InhibitEX Buffer was added to each stool sample and Vortex continuously for 1 min until the stool sample was thoroughly homogenized to ensure maximum DNA concentration in the final eluate. The homogenized sample was heated in a water bath of 70°C for 5 min and vortexed for 15 s for uniform lysis. Samples were then centrifuged at full speed (20,000×g, 14,000 rpm) for 1 min to pellet stool particles. Step 2: 25 µL proteinase K was pipetted into a new 2 mL microcentrifuge tube and 600 µL supernatant from step 1 was pipetted into the 2 mL microcentrifuge tube containing proteinase K. We Added 600 µL Buffer AL and vortex for 15 s to form a homogeneous solution which was Incubated at 70°C for 10 min and centrifugated briefly to remove drops within the tube lid. 600 µL of ethanol (96%–100%) was added to the lysate, and mix by vertexing, followed by a brief Centrifugation to remove drops from the inside of the tube lid. 600 µL of the lysate above was transferred into the QIAamp spin column and centrifuged at full speed (20,000×g, 14,000 rpm) for 1 min. The QIAamp spin column was placed in a new 2 mL collection tube, and the old tube containing the filtrate was discarded. The QIAamp spin column was carefully opened, 500 µL Buffer AW1 was added and centrifuge at full speed for 1 min. The QIAamp spin column was transferred to a new 2 mL collection tube, and old tube containing the filtrate was discarded. The QIAamp spin was carefully opened again, and 500 µL Buffer AW2 was added, and centrifuged at full speed for 3 min. The QIAamp spin column was placed in a new 2 mL collection tube and discard the old collection tube with the filtrate. Centrifuge at full speed for 3 min to eliminate the chance of possible Buffer AW2 carryover. Lastly, The QIAamp spin column was transferred into a new, labeled 1.5 mL microcentrifuge tube and 200 µL Buffer ATE was pipetted directly onto the QIAamp membrane, incubated for 1 min at room temperature, then centrifuged at full speed for 1 min to elute DNA which is then later checked using the Spectrophotometer.
For the quantification of fecal microflora such as Lactobacillus spp., Bifidobacterium spp., Clostridium spp., and E. coli., in our experiment, 1× universal SsoAdvanced™ universal SYBR® Green Supermix, 2.5 ng/μL of each forward and reverse primers, and 10 ng of DNA was added in a 10 μL volume [18,19]. Enzyme activation was achieved at the cycling parameters at 95°C followed by 40 cycles of melting at 95°C for 15 s; annealing for the specified times and temperatures according to each primer (Bioneer) and the SYBR thermal cycling protocol carried out with extension at 72°C for a specified time according to each primer, with beta actin (β-actin) as the house keeping gene (Table 3). At 72°C, the SYBR green fluorescent signals were recorded and known bacterial species were serially diluted 10-fold before being used to create PCR results. The qPCR Rotor-Gene Qiagen 2plex program with Serial Number 0312272, Corbett Research (Corbett Life Science Qiagen 2008) was used for the DNA quantification.
The ginsenoside concentration in egg yolk was measured using five egg samples for each treatment groups. 2 g of dry matter sample was extracted 3 times into 20 mL methanol while at 60°C constant temperature in a water bath at 3-hour intervals. For the extracted sample, the solvent was removed under reduced pressure under a condition not exceeding 40°C using a reflux concentrator, and the remaining residue was dissolved in 5 mL of distilled water. The dissolved residue was transferred to a separatory funnel, and the layers were separated with 50 mL of chloroform to remove non-polar components such as fat, and the organic solvent layer. At this time, the remaining residue was washed three times with 50 mL of ethyl ether. The ginsenoside layer dissolved by the aqueous layer was extracted using saturated n- butanol, and the solvent was removed under reduced pressure by observing 40°C in an evaporator. It was tested gravimetrically to determine the total ginsenoside content [13].
Data generated in the present study were subjected to statistical analysis system [20] using the general linear model (GLM) procedure in a randomized complete block design. When significant differences were identified among treatment means, they were separated using Tukey’s Honest Significant Difference test. Probability values of < 0.05 were considered significant, and cages containing six laying hens per replicate was the experimental unit.
RESULTS
The effect of WG on laying performance is shown in Table 4. The supplementation of WG showed no significant difference in ADFI and body weight (BW) from phase 1 to phase 3 across all treatments. Supplementation of WG showed no significant difference in HDEP in phase 1 but showed a tendency towards significance in phase 2 and was significantly increased (p < 0.05) in phase 3 at WG2 compared with the CON. The addition of WG had no significant effect on AEW in phase 1 and phase 2 across all treatments, but AEW was significantly increased (p < 0.05) in phase 3 at WG2 compared with the CON. The effect of WG supplementation showed no significant difference in EM across all treatments in phase 1 but showed a tendency towards significance in phase 2 and was significantly increased (p < 0.05) in phase 3 at WG2 compared with the CON. FCR was not significant in phase 1 and phase 2 of our experiment across all treatments when WG was supplemented, but it was significantly decreased (p < 0.05) in phase 3 at WG2 compared with the CON.
The effect of WG on the egg quality of layer hen is shown in Table 5. There was no significant difference in Haugh unit, yolk color, yolk weight, albumin weight, yolk percentage, albumin percentage, eggshell thickness, and hardness across all treatments from phase 1 to phase 3.
The effect of WG in the feed for laying hens on the ginsenoside content in egg yolk is shown in Table 6. There was no significant difference in phase 1 across all treatments when WG was supplemented, but the difference tended towards significant in phase 2, and there was a significant increase (p < 0.05) in the ginsenoside content in phase 3 at WG2 compared with the CON.
| Treatments1) | CON | WG1 | WG2 | SEM | p-value |
|---|---|---|---|---|---|
| Ginsenosides (mg/g) | |||||
| 4 weeks | 1.17 | 1.22 | 1.24 | 0.039 | 0.265 |
| 8 weeks | 1.22 | 1.26 | 1.34 | 0.045 | 0.084 |
| 12 weeks | 1.19b | 1.31b | 1.48a | 0.056 | 0.002 |
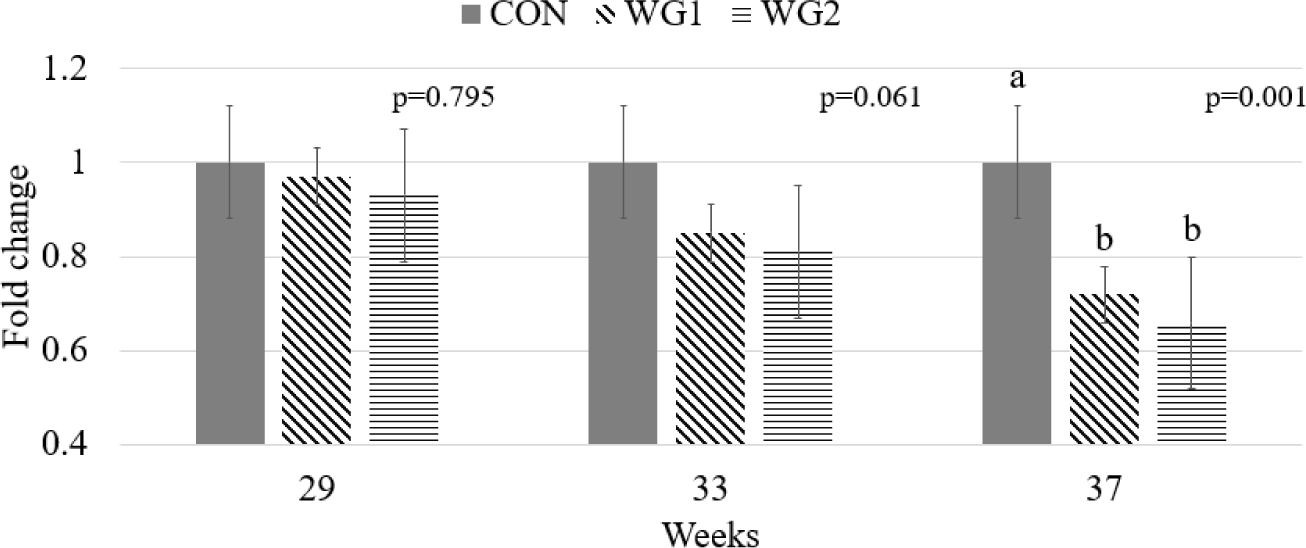
The effect of WG on the relative expression of TNF-α is shown in Fig. 1. There was no significant difference in phase 1 across all treatments when WG was added. However, phase 2 showed a tendency towards significance, and phase 3 was significantly decreased (p < 0.05) in hens on the WG-supplemented diets compared with the CON.
The effect of WG on the fecal microflora DNA of laying hens is shown in Table 7. The supplementation of WG had no significant effect on Clostridium spp. and Bifidobacterium spp. from phase 1 to phase 3 across all treatments. Lactobacillus spp. had no significant difference in phase 1 but was significantly increased (p < 0.05) in phase 2 and phase 3 at WG2 compared with the CON. Lastly, E. coli showed no significant difference in phase 1 and phase 2 but was significantly decreased (p < 0.05) in phase 3 at WG2 compared with the CON.
DISCUSSION
Ginsenosides are the primary components of ginseng and positively affect reproductive organs and other tissues [21,22]. The supplementation of WG at WG2 in the current study improved FCR, HDEP, AEW, EM, TNF-a, yolk ginsenoside content, and microflora DNA in the last phase of our experiment but had no positive effect on ADFI, BW, and egg quality. Herbs and their extracts may help the digestive system by promoting the activity of the digestive enzymes in the gastric epithelium [23]. We propose this as the reason behind the amelioration of FCR in our study. However, the bioactive contents of ginseng, such as saponin, are typically associated with a bitter taste and unpleasant sensation, which might be the reason behind the indifference in ADFI and BW across all treatments.
The increased laying performance in hens on the WG-supplemented diets in our research agrees with the study of Kang et al. and Chung et al. [24,25], where the inclusion of fermented WG by-products enhanced the laying performance of layer hens. The enhanced laying performance can be attributed to the stimulating effect of ginsenosides on oocyte meiotic maturation proliferation through the cumulus cells. The ovary of a chicken accommodates a variety of hierarchical follicles according to their maturity. One of the crucial steps in the formation of follicles related to the laying performance is the selection of small yellow follicles as pre-ovulatory follicles from a group of follicles of comparable sizes [26]. This can be further illustrated using the study by Tan et al. [27] that demonstrated that the administration of ginsenoside significantly increased the number of granular cells, a major component of the ovarian follicle, in laying hens, leading to a more cuboidal shape of the granular cells in hens on the ginsenoside-supplemented diet. This might be the reason behind the increase in laying performance in our study.
In addition, the chicken gastrointestinal pathway is susceptible to E. coli, widely distributed in their excretes, making colibacillosis one of the most widespread diseases affecting the layer and poultry sector [28,29]. Commercial layer and breeder hens with this syndrome produce fewer eggs due to exudations in the peritoneal cavity due to the inflammatory cells covering the surfaces of several organs, such as the oviduct, ovary, and intestine [30]. Additionally, the underdeveloped immune system of young layers, coupled with their high nutrient requirement, makes them more vulnerable to infections than the matured layers [31,32], which can also be the reason for the indifferent results in the early stages of our experiment. Therefore, we saw the effect of WG on mitigating the negative effect of E. coli and the modulation of inflammatory cytokine in the later phase of our study. This agrees with the study of Bi et al. [33], in which ginsenoside was able to lessen the stress caused by E. coli in broiler chickens. They explained that the impact was associated with the mammalian target of rapamycin, Hemeoxygenase-1, and superoxide dismutase overexpression, resistance against oxidative damage, and inflammatory suppression. We hypothesized from our results that the amelioration of the ginsenoside level of egg yolk in the current study could be linked to the improvement in the TNF-α and microflora DNA expression, leading to a reduction in the presence of negative bacteria E. coli and an increase in the positive Lactobacillus in the later phase of our experiment. In detail, ginsenoside’s amphiphilic and hydrophilic carbohydrate nature enables their adhesion into membranes which also entails lamella collections of phospholipids with amphiphilic ends, allowing them to interact with the interfacial region membranes made up of multiple glycolipids and glycoprotein of the yolks allowing the steroid portion of the saponins to engage with sterol membrane. This enable the saponin osidic component to promote the development of intramolecular hydrogen bonding [13,34]. Furthermore, Fukuda et al. [34,35] revealed that ginsenosides shows a distinctive agglutinability to egg yolk vesicles which is phosphatidylcholine in nature, with three-dimensional structure serving as the final determining factor of its ability to agglomerate with other lipids to form a peripheral membrane domain [36–38]. We hypothesize this leads to an increase in O-glycosidic oligosaccharides enabling a better gut and immune system.
Lastly, our research showed no significant differences in egg qualities, such as haugh unit, yolk color, yolk weight, albumin weight, yolk percentage, albumin percentage, eggshell thickness, and hardness across all treatments and phases. This is contrary to the study of Jang et al. [39], in which red ginseng enhanced egg quality. We propose that such discrepancies may be attributed to different ginseng strains, sources, or preparation techniques used in the studies.
CONCLUSION
In conclusion, the supplementation of WG in diets led to an increase in yolk ginsenoside content and reduce TNF-α, ultimately leading to an improved microflora DNA quality and laying performances, such as HDEP, AEW, EM, and FCR in hens on the WG2-supplemented diets. Therefore, we recommend the use of 0.5 % WG per basal diet for improved laying performance in layer hens.