
INTRODUCTION
As the second limiting amino acid (AA) in corn and soybean meal diets for poultry, lysine (Lys) is commonly utilized as a reference AA to establish the ideal AA ratios [1,2]. Lys plays a central role in supporting the growth performance of poultry by promoting nutrient utilization and muscle development as well as protein synthesis and production of enzymes, hormones, and antibodies [3,4]. Accordingly, determining the optimal dietary Lys content is necessary for achieving efficient duck production.
Based on observations in the marketing of various poultry species, it is evident that the industry has made significant strides in the development of cut and processed duck products. This progress can be attributed to the implementation of genetic selection and advancements in duck management, particularly in the realm of nutrition. As a result, the meat yield of ducks has experienced a notable increase, while carcass fatness has concurrently decreased [5,6]. Hence, it becomes imperative to formulate updated nutrient requirements to meet the evolving demands of genetic enhancements in meat-type ducks.
Numerous investigations have been carried out to ascertain the Lys requirement for White Pekin ducks [7–9]. However, there is the suggestion that using standardized ileal digestible (SID) AA could offer a more accurate means of determining these requirements in animals, as it accounts for the bioavailability of AA from various feed ingredients [10,11]. This method assesses the disappearance of AA in the small intestine, providing a more reliable indicator of AA digestibility without disrupting the hindgut [12]. Despite this rationale, limited attention has been given to recent studies that focus on estimating the SID Lys requirements during the starter period (up to 21 days of age) in White Pekin ducks. Moreover, a range of regression models, such as the linear broken line and the quadratic broken line can be effectively employed to estimate the digestible Lys requirements for ducks, as demonstrated by the research conducted by [13]. The utilization of distinct estimation models provides diverse dietary Lys requirements, facilitating the determination of optimal nutritional Lys concentrations for enhancing animal breeding practices [14,15]. Therefore, this study aims to determine the SID Lys requirement for ducks from hatch to 21 days of age, utilizing both the linear broken-line and quadratic line models.
MATERIALS AND METHODS
The Animal Ethics Committee of Chungnam National University, Daejeon, Korea, approved the protocols used in this experiment (approval number: 202109A-CNU-114).
The experimental diets (detailed in Table 1) comprised six variations with progressively increasing SID Lys concentrations. SID values for AA in corn, soybean meal, and corn distillers’ dried grains with solubles (DDGS) were sourced from a prior investigation [16]. These dietary formulations encompassed SID Lys concentrations ranging from 0.72% to 1.12%, incremented by every 8 points. Each experimental diet was meticulously crafted to either meet or surpass recommended specifications [17], except for Lys, which was adjusted to align with the requirements of ducklings at 3 weeks of age. Indispensable AA concentrations, excluding Lys, were calibrated based on ideal AA ratios to avert deficiencies. The experimental diets were provided in crumble form.
Provided per kilogram of diet: vitamin A, 12,000 IU; vitamin D3, 2,500 IU; vitamin E, 30 IU; vitamin K3, 3 mg; D-pantothenic acid, 15 mg; nicotinic acid, 40 mg; choline, 400 mg; and vitamin B12, 12 μg; Fe, 90 mg from iron sulfate; Cu, 8.8 mg from copper sulfate; Zn, 100 mg from zinc oxide; Mn, 54 mg from manganese oxide; I, 0.35 mg from potassium iodine; Se, 0.30 mg from sodium selenite.
The experiment was carried out in two consecutive periods, with 192 birds in each period, within the same research facility due to space constraints. Consistent procedures and environmental conditions were maintained throughout. The experiment was conducted using 384 male White Pekin ducklings from hatch to 3 weeks of age. One-day-old male White Pekin ducklings were obtained from a local hatchery (Charmfre, Jincheon, Korea) for the experiment. Upon arrival, the ducklings were weighed and randomly allocated to one of the six dietary treatments with varying digestible Lys levels. Each pen, measuring 1.7 m × 1.3 m × 1.0 m, housed eight birds with a mean body weight (BW) of 53.05 ± 0.201 g (mean ± SEM). The floor pens were lined with rice husk as litter, following the recommendation of a previous study we conducted [18], and each pen was equipped with tree nipple drinkers and a feeder. The ducklings had ad-libitum access to the experimental diets and fresh water for 21 days. Continuous lighting was provided for 24 h, and the ambient temperature was maintained at 30-32°C for the first week, gradually decreasing to 25°C until 21 days of age.
The initial BW of the birds was recorded upon arrival, and subsequent BW and feed consumption were measured weekly (on days 7, 14, and 21) throughout the experiment. Based on these measurements, the average daily gain (ADG), mortality-corrected average daily feed intake (ADFI), and feed conversion ratio (FCR) were calculated for each cage during each respective week. The AA composition of the experimental diets was determined using standard procedures (AOAC method 982.30 E) [19]. The analyzed AA content of the experimental diets is shown in Table 2.
The collected data were analyzed according to a completely randomized design using the general linear model procedure for the one-way ANOVA using SPSS software (Version 26, IBM SPSS, IBM, Armonk, New York, USA). Each pen served as the experimental unit for all growth performance measurements. Orthogonal polynomial contrasts were conducted to assess the significance of linear or quadratic effects of SID Lys levels on all measurements. When significant treatment effects were observed (p < 0.05), means were separated using Tukey’s multiple range test in SPSS software. Linear-plateau and quadratic-plateau regression analysis, performed with the Nutritional Responses Model version 1.3 [13], were used to estimate the SID Lys requirements.
RESULTS
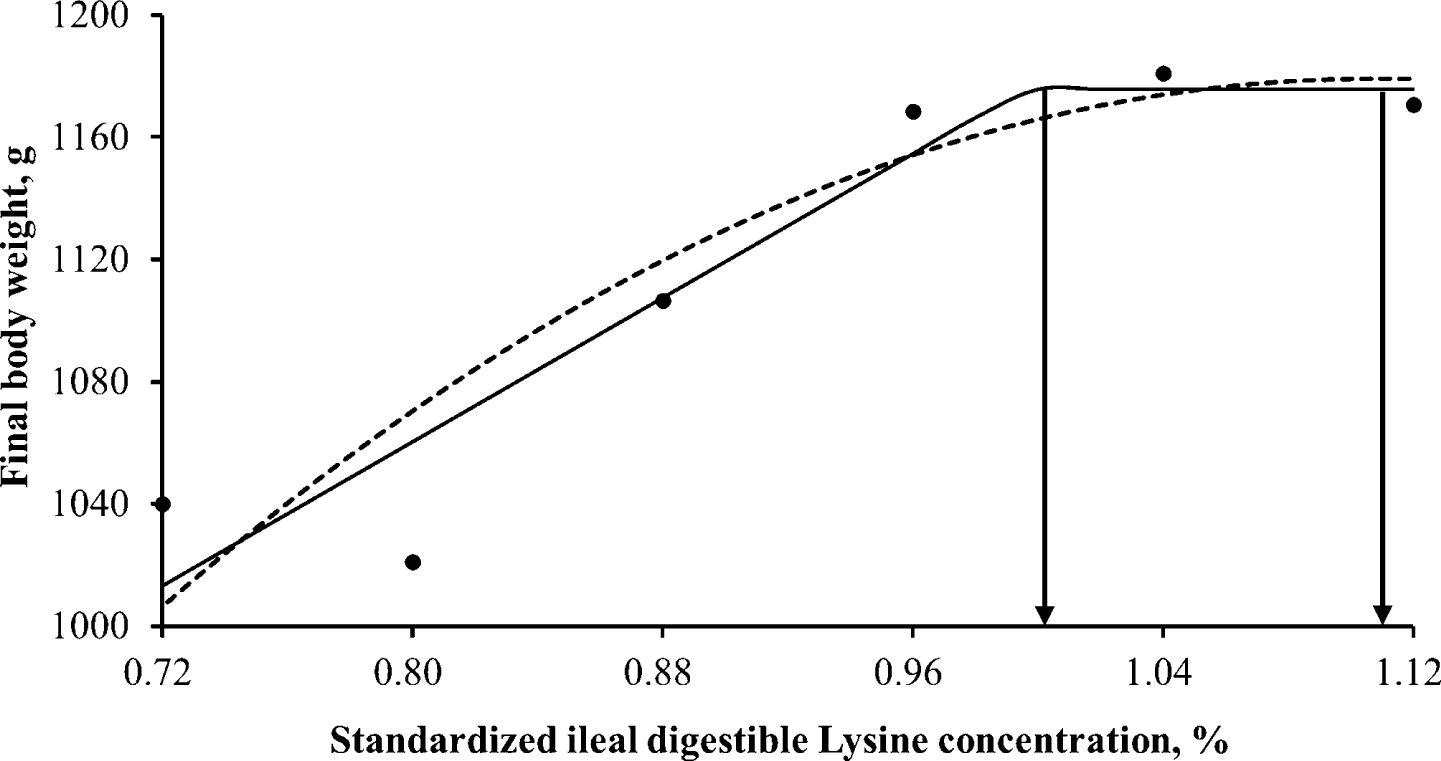
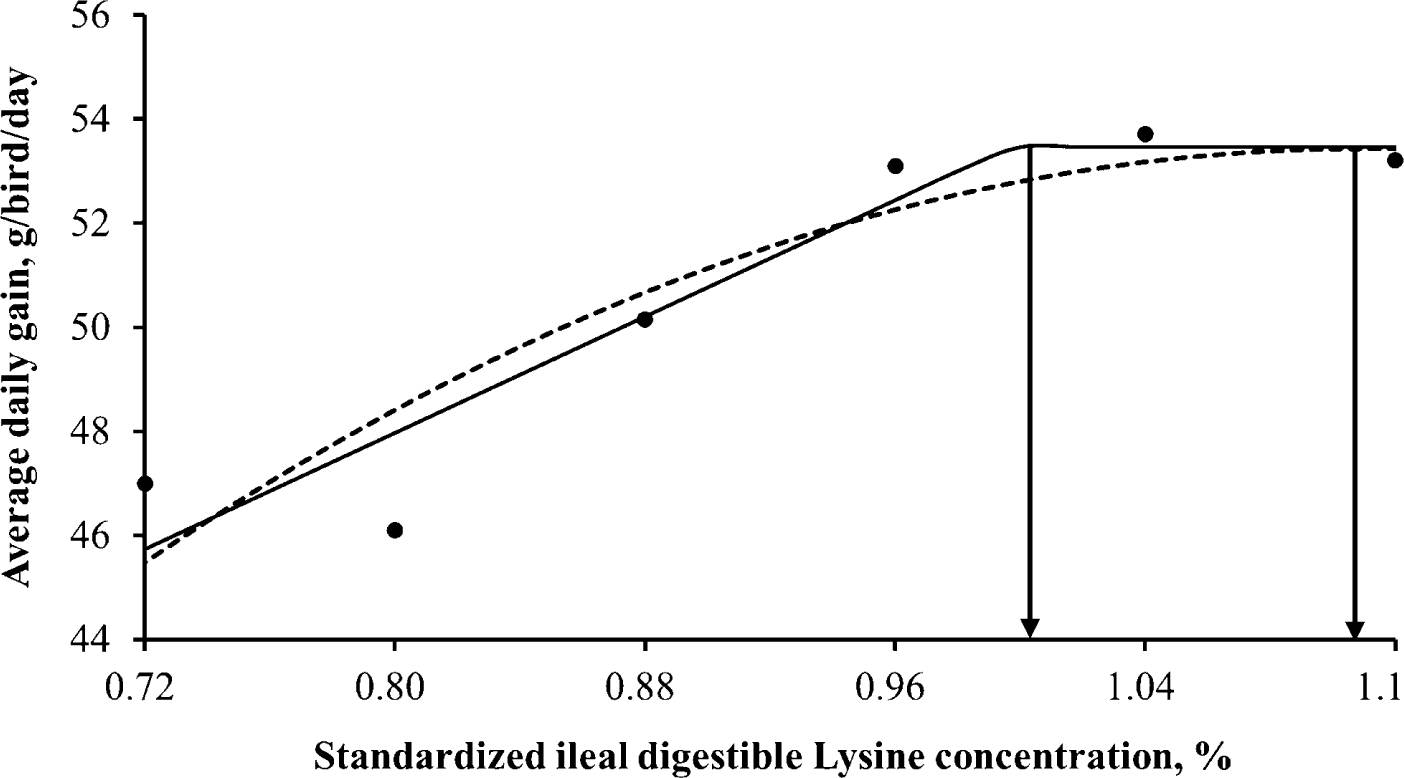
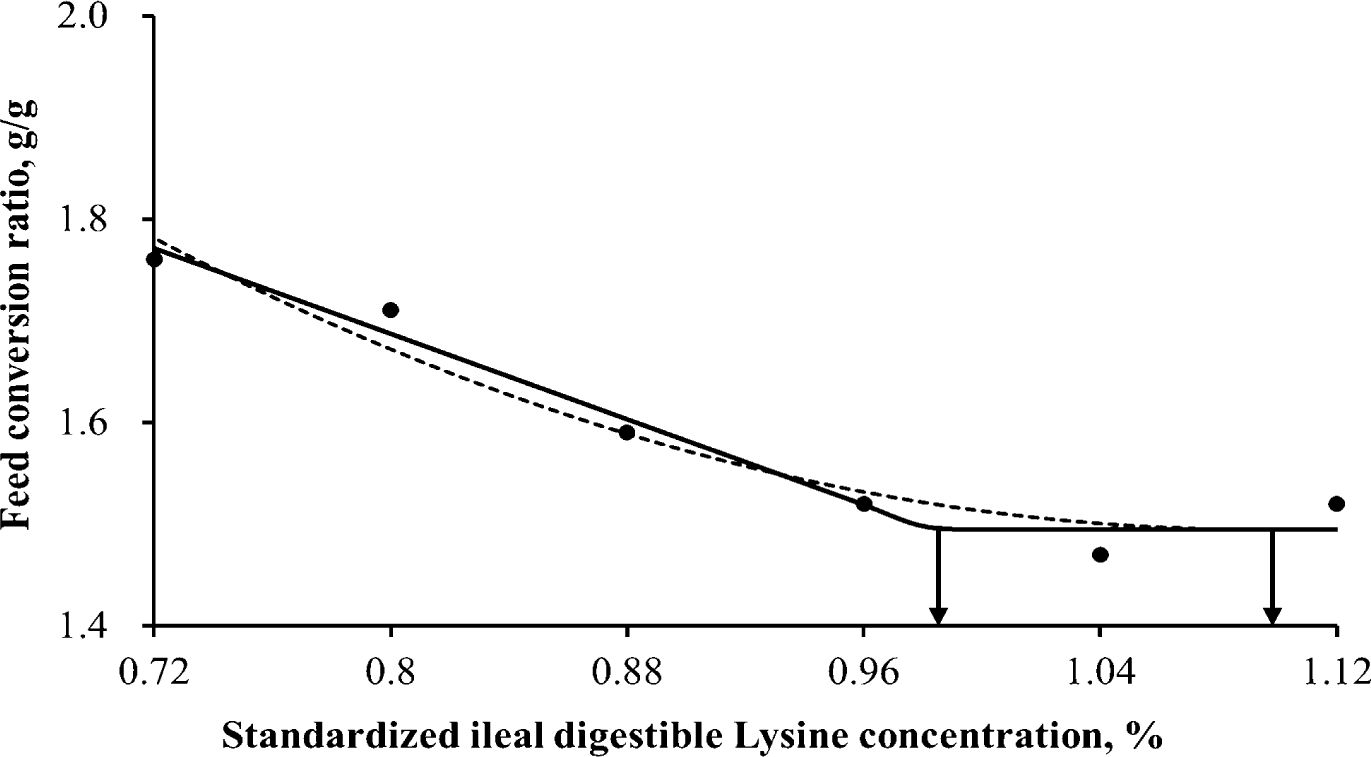
Throughout the entire 3 weeks experiment, the ducklings remained in good health and performed well. Different levels of dietary SID Lys ranging from 0.72% to 1.12% across 6 treatments in the experimental diets resulted in notable enhancements (p < 0.001) in BW, ADG, and feed efficiency for 3 weeks after hatching, with linear and quadratic manners (Table 3). SID Lys requirements for White Pekin ducks during the 3 weeks after hatch were estimated (Table 4) using two different response models. When data were analyzed using a linear-plateau model, the estimated requirements were 1.00% and 1.00% for maximum final BW and ADG, respectively, while the requirement for minimum FCR was 0.98% (Figs. 1, 2, and 3). On the other hand, the quadratic-plateau model yielded estimates of 1.11% and 1.11% for maximum final BW and ADG respectively, and 1.10% for minimum FCR (Figs. 1, 2, and 3). By averaging the values obtained from both response models, the recommended SID Lys requirements for White Pekin ducks during the 2 wk after hatching were determined as 1.05%, 1.05%, and 1.04% for maximum final BW, ADG, and minimum FCR, respectively.
DISCUSSION
Our study aimed to assess the SID Lys requirement for achieving the ideal growth performance during 3 wk after the hatch in White Pekin ducks. Although numerous studies have investigated the SID Lys values in feed formulations to determine the ideal Lys requirements for broiler chickens [15,20,21], there exists a notable scarcity of published data specific to White Pekin ducks in this regard.
A comparative analysis of AA digestibility was conducted between broiler chickens and Pekin ducks [22]. The findings strongly indicate that utilizing values derived from feedstuffs formulated for broiler chickens should be avoided when formulating diets specifically for ducks. This recommendation is primarily attributed to the higher levels of basal endogenous AA losses observed in ducks in comparison to broiler chickens. As a result, it is imperative to consider these contrasting factors in diet formulation to ensure the best nutrient utilization for ducks. For these reasons, the formulation of experimental diets was based on the consideration of SID AA content in this study. The utilization of SID AA content as a measure is considered more precise compared to total or dietary AA content, as it reflects the nutrient availability for birds [23]. The SID AA values for the diets were determined by incorporating digestible coefficients specific to ducks [12], as well as the total AA content of the ingredients.
The present study observed increasing the SID Lys level had a non-linear impact on various performance parameters, including BW, ADG, ADFI, and feed efficiency. This finding aligns with previous research by Bons et al. [7] and Xie et al. [9], which also demonstrated a non-linear improvement in performance indicators of male White Pekin ducks with increasing Lys levels.
Precise identification of an appropriate statistical model holds paramount importance in accurately estimating nutrient requirements, as the choice of model can significantly influence the derived requirement values [24]. The variation in nutrient recommendations can arise due to the application of different estimation models, which is a common practice observed in similar experiments [13]. This highlights the need for careful consideration when selecting an appropriate model to derive accurate and consistent nutrient requirement estimations. The linear plateau model, although it may exhibit a satisfactory statistical fit, has a tendency to underestimate the optimal nutrient requirements of the animal groups studied due to their failure to consider the physiological variances present within the population [25]. Conversely, the quadratic plateau model estimates higher nutritional requirements compared to the linear plateau model [26]. Therefore, a combined approach, averaging the results of the linear plateau and quadratic plateau models, was employed to estimate Lys requirements for White Pekin ducks during the critical 21 d period after hatching.
In the current investigation, the linear plateau and quadratic-plateau regression analyses determined that the minimum requirement of SID Lys for achieving maximum BW and ADG was determined to be 1.05%, while the minimum requirement for attaining optimal FCR was found to be 1.04%. These findings align with the study conducted by Bons et al. [7], which indicated that reaching 95% of the asymptote in ADG for White Pekin ducks occurred at a total Lys concentration of 1.17% (day 1 to 21). Additionally, for efficient FCR, male Pekin ducks required a 1.06% Lys concentration from day 1 to 21. Similarly, Xie et al. [9] reported Lys requirements of 0.84% for ADG and 0.90% for feed conversion efficiency of male White Pekin ducklings from day 7 to 21, both of which exceeded the recommendations of NRC [17]. These differences in Lys requirements can be attributed to variations in response criteria, research methodologies (including experimental diets based on digestible AAs), and the enhanced growth potential resulting from the genetic selection of Pekin ducks [27]. Furthermore, it is worth noting that the response to Lys may be affected by the concentrations of other AA in the diet [28]. Furthermore, the selection of an appropriate mathematical model can have a significant impact on the estimation process [29].
CONCLUSION
The findings of this research demonstrate that augmenting the SID Lys content has a positive impact on ADG and feed efficiency during the 3 weeks after hatching in White Pekin ducks. By employing both linear- and quadratic-plateau models, it was determined that the recommended SID Lys levels for optimal final BW, ADG, and feed efficiency in White Pekin ducks from hatch to 21 d are 1.05%, 1.05%, and 1.04%, respectively.