INTRODUCTION
Mastitis in bovine cows represents the most widespread disease within the contemporary cattle industry, leading to significant economic losses. These losses severely hinder the development of dairy farming as well as the hygiene and safety of human food [1]. Currently, the global treatment for dairy cow mastitis predominantly involves antibiotics, whose prolonged and extensive use may result in internal flora dysfunction, organism resistance, and antibiotic residues in milk [2]. Consequently, the quest for new alternative therapies and treatments is a challenge that must be addressed by veterinary practitioners and researchers collaboratively. Traditional Chinese herbs offer several benefits over antibiotics, including reduced side effects, a lower likelihood of bacterial resistance, minimal toxicity, negligible residue, and no dependency. Many traditional Chinese medicines or traditional Chinese medicine extracts have been shown to be effective in the treatment of bovine mastitis (BM). Wang J used the flavonoid compound Morin isolated from Moraceae Chinese herbal medicine to act on lipopolysaccharide (LPS)-induced bovine mammary epithelial cells (BMECs). He found that Morin could significantly reduce the gene expression of inflammatory factors and inhibit the activation of NF-κB and MAPK signaling pathways, thereby achieving the anti-inflammatory effect of treating cow mastitis [3]. Shen et al. used S. haemolyticus to induce the inflammatory response of BMECs. He found that the mRNA and protein levels of inflammatory factors in BMECs were significantly reduced after paeoniflorin (PF) treatment, and the activation of TLR2 and NF-κB signaling pathway-related genes and protein expression were inhibited, indicating that PF is expected to be a potential drug for the treatment of BM [4]. In addition, Safia A found the bacteriostatic effect of Chinese herbal medicine. He tested the bacteriostatic effect of squaw mint, catnip, and lemon balm on the pathogenic bacteria of BM. The results showed that essential oil and some herbal plant extract had significant bacteriostatic activity against S.aureus and Escherichia coli, and pointed out that lemon balm and peppermint oil could be used as alternative methods for the treatment of mastitis [5]. These research results show that traditional Chinese medicine has great potential in the treatment of mastitis. Furthermore, the treatment of mastitis with Chinese herbal medicine focuses on enhancing blood circulation in the udder, clearing blocked vessels, relieving stagnation, and boosting the body’s immunity, thereby providing both symptomatic relief and addressing the root cause of the disease. This approach offers fresh perspectives in the search for alternative therapies.
Danggui Buxue decoction (DBD) was obtained from Li Dongyuan’s Discussion of Internal and External Injuries and is known as a classic formula for tonifying qi and generating blood [6]. DBD was made by decocting Astragalus and Angelicae at a ratio of 5:1 in two volumes of water and filtering and concentrating the mixture [7]. In the formula, Astragalus can strengthen the spleen, act as a diuretic, and detoxify qi, the main treatment for qi and blood deficiency. Angelica sinensis (AR) can enhance blood production, prevent thrombosis, relieve pain, and treat blood deficiency. Angelica and Astragalus complement each other, facilitating the tonification of qi and the generation of blood. Several studies have shown that DBD has biological functions, such as anticancer, antitumor, antioxidant, antifibrosis, accelerated hematopoiesis, and immunomodulatory activities [8–14]. However, the efficacy of DBD on mastitis in dairy cows has not been reported.
BM is identified by a diverse array of pathogenic factors and a complex pathogenesis. The expression of inflammatory factors such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6, among others, is elevated in the mastitis tissues of dairy cows compared to healthy tissues [15]. Subsequent research has indicated a close association between cell damage, oxidative stress, and the progression of mastitis. Upon cellular inflammation, an increase in lactate dehydrogenase (LDH) release is observed alongside the upregulation of BNBD5, GM-CSF, CCL2, and GADD-45α expression. LDH serves as a crucial indicator of the extent of cellular damage; GM-CSF and CCL2 facilitate the convergence of macrophages to damage sites and modulate an organism’s immune response; BNBD5 represents a potential local defense gene for breast tissue; and GADD-45α, which is inducible by various damage factors, is a DNA damage-responsive gene. Additionally, LPS-induced cellular inflammation may lead to an increase in reactive oxygen species (ROS) and the upregulation of COX-2, PPARγ, and iNOS expression [16,17]. COX-2, which is typically not expressed under resting conditions, can be rapidly upregulated during inflammation, while prolonged activation of iNOS may result in excessive NO production and oxidative stress, causing tissue damage [18,19]. Total antioxidant capacit (T-AOC), glutathione (GSH), superoxide dismutase (SOD), malondialdehyde (MDA) and NO can reflect the body’s oxidation and antioxidant [20]. It has been noted that in mastitis induced by gram-negative bacteria, TLR4 specifically recognizes LPS, mediates MyD88 signaling, activates NF-κB, initiates the transcription of related factors, and promotes cytokine release [21,22]. These findings highlight key molecules as potential therapeutic targets for bovine mastitis. Research has demonstrated the modulatory effects of DBD on several of the aforementioned factors. For instance, Li et al. reported that DBD treatment diminished the production of IL-1β, IL-6, IL-10, and TNF-α in a mouse model of inflammatory bowel disease (IBD) induced by dextrose sodium sulfate (DSS), effectively improving inflammation and pathological conditions [23]. Zhang Yan’s study suggested that DBD, in combination with conventional therapy, could lower TNF-α and IL-6 levels in diabetic nephropathy mice, mitigating the inflammatory response [24]. Research by Liu et al. indicated that DBD notably decreased LDH levels in mice with cardiac injury and downregulated NF-κB protein expression in cardiac tissues [25]. Gong et al. identified the main components of the DBD and formulated an herbal soup, “DBTΔfa,” which was found to decrease ROS formation and enhance the transcriptional activity of antioxidant genes in H9C2 cells [26]. Wang et al. proposed that the therapeutic effects of DBD on rats with bleomycin-induced pulmonary fibrosis might occur through the inhibition of the TLR4/NLRP3 signaling pathway [27]. Additionally, the inhibition of NF-κB activity by DBD was shown to reduce inflammation-related damage [28]. The experimental findings of Gong et al. demonstrated that the modulation of the NF-κB signaling pathway by DBD has immunomodulatory effects [29]. This finding suggests the potential of DBD to treat mastitis by inhibiting the production of these factors.
The aim of this study was to investigate the efficacy and mechanism of action of DBD in treating bovine mastitis and to fill this research gap in the field of DBD treatment for bovine mastitis. We used LPS to induce inflammation in BMECs, which was used to establish an in vitro cellular model of bovine mastitis, to evaluate the protective effect of DBD on BMECs in terms of inflammation, injury and oxidative stress to conduct preliminary studies on the molecular mechanisms involved. Our experimental results provide basic information for further research on the mechanism of action of DBD in mastitis in dairy cows and provide a reference for the use of DBD in the prevention and treatment of bovine mastitis.
MATERIALS AND METHODS
BMECs were preserved at the Veterinary Obstetrics Lab of Gansu Agricultural University (Lanzhou, China). RPMI 1640 medium was obtained from Gibco (C11875500BT). LPS (From Escherichia coli O55:B5) was obtained from Solarbio (L8880). Astragalus membranaceus (ASR) and AR were obtained from the Yellow River Herbal Market. TRIzol was obtained from Ambion (15596–026). An Evo M-MLV RT Kit with gDNA Clean for quantitative polymerase chain reaction (qPCR) was obtained from Accurate Biotechnology (AG11728). 2×SYBR Green qPCR Master Mix was obtained from Selleck (B21203). A bovine LDH Cytotoxicity Assay Kit was obtained from Sino Best Biological (YX-22476B). The CCK-8 (C0042), ROS assay kit (S0033S) and NO assay kit (S0021S) were obtained from Beyotime. The T-AOC assay kit (BC1315) was obtained from Solarbio. The GSH assay kit (A006-2-1), SOD assay kit (A001-3) and MDA assay kit (A003-1) were obtained from Nanjing Jiancheng Bioengineering Institute. The GS-Prestained Protein Ladder was obtained from Genesand (PM901). Cell lysis buffer (P0013) and QuickBlock™ Blocking Buffer (P0252) were obtained from Beyotime. Rabbit anti-IL-1β antibody (bs-20449R) (1:2000), rabbit anti-IL-8 antibody (bs-0780R) (1:2000) and rabbit anti-NF-κB p65 antibody (bs-0465R) (1:1000) were obtained from Bioss. A rabbit anti-IL-6 antibody (21865–1-AP) (1:1000) was obtained from Proteintech. Rabbit anti-TNF-α antibody (AF-7014) (1:2000) was obtained from Affinity Biosciences. Rabbit Anti-GM-CSF antibody (CPA2949) (1:1000), Rabbit Anti-CCL2 antibody (CQA3687) (1:1000), Rabbit Anti-GADD-45α antibody (CQA1970) (1:1000), Rabbit Anti-TLR4 antibody (CPA2171) (1:1000), Rabbit Anti-NF-κB p50 antibody (CPA4479) (1:1000), Mouse Anti-β-actin antibody (CPA9100) (1:10000) was from Cohesion Bio. Horseradish peroxidase (HRP)* goat anti-mouse IgG (H+L) (RS0001) (1:10000) and HRP* goat anti-rabbit IgG (H+L) (RS0002) (1:10000) were purchased from ImmunoWay Biotechnology.
DBD was extracted from AR and ASR at a ratio of 5:1, and these herbs were obtained from Gansu Province. The herbal liquid was soaked in distilled water (1:8; v/w) for 30 min and boiled in water for 1 h, after which the residue from the first extraction was boiled in water (1:6; v/w) for 1 h. To prepare the DBD solution, after being mixed twice, the solution was subjected to concentration using a rotary evaporator [30]. Subsequently, the DBD solution was concentrated using a rotary evaporator at 60°C and 60 rpm (approximately 0.5×g), and the concentrated drug solution was freeze-dried at −50°C to obtain the DBD powder. Before use, the DBD powder was dissolved in RPMI 1640, and the DBD medium was heated at 50°C, sonicated for 15 min, and filtered through a 0.22 μm filter. The desired concentrations were prepared as needed.
The cells were cultured at 37°C in a 5% CO2 cell culture incubator, 0.25% trypsin was used for digestion, RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS) was used for passaging culture, and RPMI 1640 medium without FBS was used for preparation of the required drugs.
A total of 100 μL of the BMECs suspension at a concentration of 5 × 104 cells/mL was inoculated into a 96-well plate, and subsequent studies were performed after the cells had grown to confluency. BMECs were grown in 96-well cell culture plates, 100 µL of DBD at final concentrations of 10, 25, 50, 100, 250, 500, 1,000, 2,500, 5,000, 10,000, 25,000, 50,000, and 100,000 µg/mL was added to the DBD group, and LPS was added to the LPS group at final concentrations of 0.1, 1, and 10 µg/mL. The culture plate was incubated at 37°C for 24 h. A total of 10 µL of CCK-8 reagent was added to each well 4 h before the end of the incubation, after which the cells were incubated. After the incubation was completed, the optical density (OD)450 of each well was measured with an enzyme labeler, and the cell viability was calculated (unit: %). When the cell viability was less than 100%, the cell inhibition rate (IR) was calculated as follows: IR = (100-cell viability) × 100%, and the IR ≤ 10% criterion was used to establish the upper limit of the supplied concentration of DBD medium.
2 mL of BMECs cell suspension at a concentration of 5 × 105 cells/mL was inoculated into a 6-well plate, and subsequent studies were performed after the cells had grown to the wall. LPS is a component of the cell wall of gram-negative bacteria. LPS induces an inflammatory response in cells, and LPS-treated cell models are widely used in inflammation studies. To establish an in vitro cellular model of bovine mastitis, we stimulated BMECs for 24 h with LPS at final concentrations of 0.1, 1, and 10 µg/mL. The mRNA and protein of the cells were collected, and the expression of TNF-α, IL-1β, IL-6, and IL-8 at the transcriptional and protein levels was analyzed via qPCR and western blot (WB).
To observe the effects of DBD on the expression of TNF-α, IL-1β, IL-6, IL-8, BNBD5, GM-CSF, CCL2, GADD-45α, COX-2, PPARγ, iNOS, TLR4, and NF-κB in LPS-stimulated BMECs, the cells were cultured for 24 h with 1 µg/mL LPS as the LPS group, and the cells were cocultured for 24 h with 100 µg/mL DBD and 1 µg/mL LPS as the LPS + DBD group. For analysis of protein and mRNA expression, the cells in 6-well plates were collected. Bovine LDH cytotoxicity and NO analyses were performed using the cell culture supernatant collected from 6-well plates. Walled cells were subjected to ROS staining.
Total RNA was extracted with TRIzol, and the A260/A280 ratio of RNA was determined to be between 1.8 and 2.0 before reverse transcription. An Evo M-MLV RT Kit with gDNA Clean was used to reverse transcribe the RNA to cDNA. The internal reference gene was GAPDH; the target genes were TNF-α, IL-1β, IL-6, IL-8, BNBD5, GM-CSF, CCL2, GADD-45α, COX-2, PPARγ, iNOS, TLR4, NF-κB p65 and NF-κB p50. The reaction mixture contained 10 µL of 2×SYBR Green qPCR Master Mix, 1 µL of each forward and reverse primer, and 1 µL of cDNA, for a total volume of 20 µL. The reaction conditions were 95°C for 600 s, 45 cycles of 95°C for 15 s, and 60°C for 45 s. A LightCycler®96 real-time detection system (Roche) was used to carry out the reaction program. The 2–ΔΔCT method was utilized to determine the relative expression of the target genes. Table 1 summarizes the sequences of the primers used in this study.
BMECs were inoculated into 6-well plates, and at the end of the treatment, an appropriate amount of cell lysis buffer was added. The lysate was centrifuged at 6,000×g for 10 min at 4°C, after which the precipitate was removed to obtain protein samples. The protein concentration was determined using a BCA protein assay kit, and an equal amount of protein was added to a protein electrophoresis gel and separated via sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS‒PAGE). The proteins were then electrotransferred onto a polyvinylidene fluoride (PVDF) membrane via wet transfer, and the resulting PVDF membrane was incubated with QuickBlock™ Blocking Buffer for 1 h at room temperature. The membranes were incubated with rabbit polyclonal antibodies against TNF-α, IL-1β, IL-6, GM-CSF, CCL2, GADD-45α, TLR4, NF-κB p50, and NF-κB p65 and mouse polyclonal antibodies against IL-8 and β-actin overnight at 4°C. After the membrane was washed, the sections were incubated with HRP* goat anti-mouse IgG (H+L) and HRP* goat anti-rabbit IgG (H+L). Immunoreactivity was observed on an Amersham ImageQuant 800 (Cytiva) after the dropwise addition of enhanced chemiluminescence (ECL) reagent to the PVDF membranes, and the gray values were analyzed using ImageJ software (National Institutes of Health).
For the LDH release assay, the cell culture fluid was centrifuged. According to the instructions, 50 µL of LDH standards at different concentrations (0, 30, 60, 120, 240, and 480 U/L) were added to the standard wells to construct a standard curve, and 10 µL of cell culture solution from different treatments and 40 µL of sample diluent were added to the sample wells. Then, 100 µL of HRP-conjugate reagent was added to each well, and the mixture was allowed to stand at 37°C for 60 min. The substance was removed, and the plate was rinsed five times with washing solution. Then, 50 µL of Chromogen Solution A and 50 µL of Chromogen Solution B were added to each well, which was incubated for 15 min at 37°C in the dark. Afterward, 50 µL of Stop Solution was added, and the OD value of each well was measured at 450 nm using an enzyme meter. A standard curve was plotted with the OD at 450 nm as the horizontal coordinate and the concentration of LDH (U/L) as the vertical coordinate, and the concentration of LDH in the sample was determined.
Cells in 6-well plates were collected and lysed by adding a proper amount of pre-cooled extract and centrifuged at 6,000×g for 10 min. The supernatant was taken to determine the protein concentration and detect T-AOC. 40 μmol/mL FeSO4 standard solution was prepared and diluted to various concentrations. Standard tube (100 μL FeSO4 solution + 100 μL reagent 2), blank tube (24 μL distilled water + 180 μL mixture) and determination tube (6 μL sample + 18 μL distilled water + 180 μL mixture) were set up. After fully mixed, 200 μL was added to the 96-well plate after 10 min, and the OD value was measured at 593 nm.
The standard dilution was used to prepare 20 μM GSH standard solution, and then 100 μL of cell protein sample was mixed with 100 μL precipitation solution. After centrifugation, the supernatant was taken for testing. 100 μL precipitation liquid was added to the blank well, 100 μL 20 μM GSH was added to the standard well, and 100 μL supernatant was added to the determination well. 100 μL buffer and 25 μL chromogenic agent were added to each well and mixed. After standing for 5 min, the OD value of 405 nm was measured, and the GSH content (mol/gprot) in each group was calculated.
The appropriate dilution ratio of cell protein was selected in the pre-experiment. According to the instructions of SOD assay kit, the control well, the control blank well, the determination well, and the determination blank well were set up. After adding the corresponding reagents to each well, mixing well, and standing at 37°C for 20 min, the OD value of each well at 450 nm was measured. The SOD IR (%) and SOD activity (U/mgprot) in each group were calculated.
According to the instructions of the MDA assay kit, a blank tube (100 μL anhydrous ethanol), standard tube (100 μL 10 nM tetraethoxypropane) and determination tube were set up. Add 4 mL of pre-prepared working solution to each tube and mix well, boil for 40 minutes, and then rinse with water to cool. The cooled samples were centrifuged at 800×g for 10 min. The supernatant obtained from 200 μL was added to a 96-well plate, and the OD value was measured at 532 nm to calculate the MDA content (nmol/mg prot) of each group.
For the NO content assay, the cell culture fluid was centrifuged. The method was carried out in accordance with the manufacturer’s instructions after all the reagents were brought to room temperature. The 96-well plate was filled with 50 µL of the test samples, 50 µL of standards at various concentrations, and 50 µL of each of the Griess Reagent I and Griess Reagent II. Within 10 min, the absorbance value at 540 nm was determined. A standard curve was plotted with the OD value at 540 nm as the horizontal coordinate and the concentration of NaNO2 (μM) as the vertical coordinate, and the final concentration of NO in the sample was obtained.
BMECs 1 mL suspension at a concentration of 5×105 cells/mL was inoculated into glass-bottomed culture dishes and treated with drugs after the cell growth density reached 80%. The cells were incubated with Rosup (50 µg/mL) at 37°C for 30 min as a positive control for ROS accumulation. Using the in situ loading probe assay, dichlorodihydrofluorescein diacetate (DCFH-DA) at a concentration of 10 µmol/L was prepared with RPMI Medium 1640 basic, and 1 mL of DCFH-DA was added to the cells, which were incubated at 37°C for 30 min. The cells were subsequently washed three times with RPMI 1640 medium to minimize residual DCFH-DA. The cells in each well were observed by fluorescence microscopy after the addition of an anti-fluorescence quencher.
RESULTS
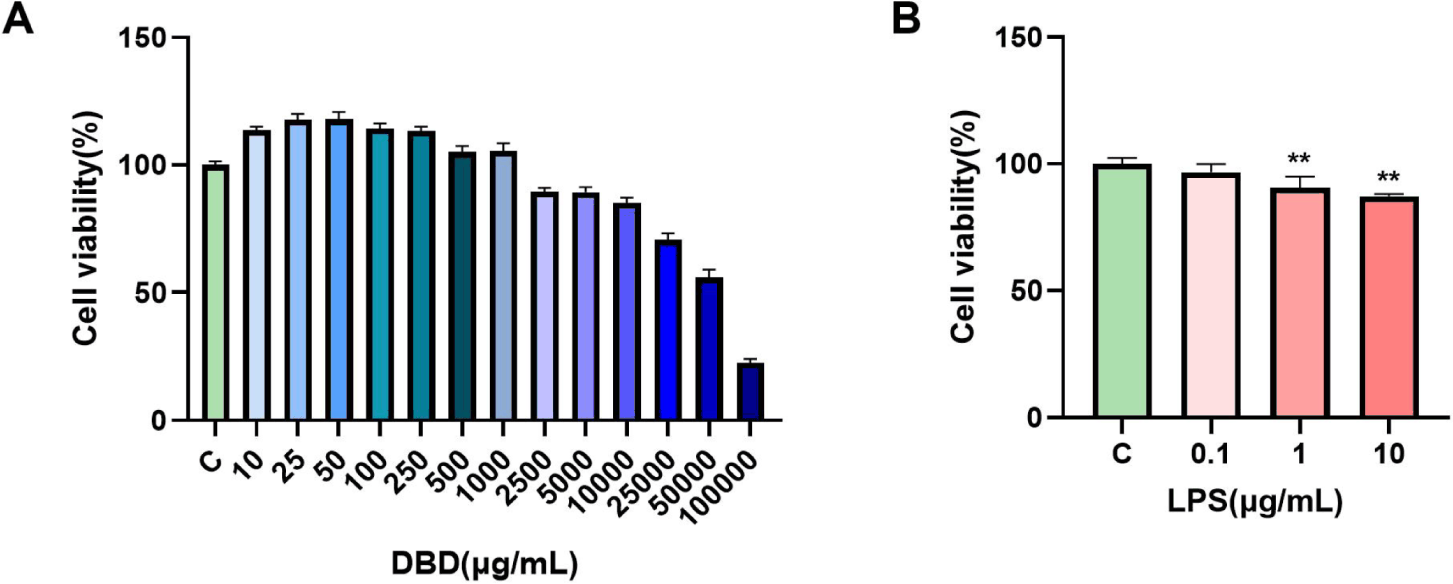
The CCK-8 assay was used to measure the cytotoxicity of DBD and LPS on the BMECs to examine their impact on their viability. These findings demonstrated that whereas high amounts of DBD reduced cell growth, low quantities of DBD promoted cell growth. The maximum concentration limit for DBD-treated cells was found to be 1000 µg/mL since the IR was greater than 10% at 2,500 µg/mL (Fig. 1A). Moreover, we discovered that LPS suppressed cell proliferation in a concentration-dependent manner and that it could significantly limit cell growth at concentrations up to 1 µg/mL (**p < 0.01) (Fig. 1B).
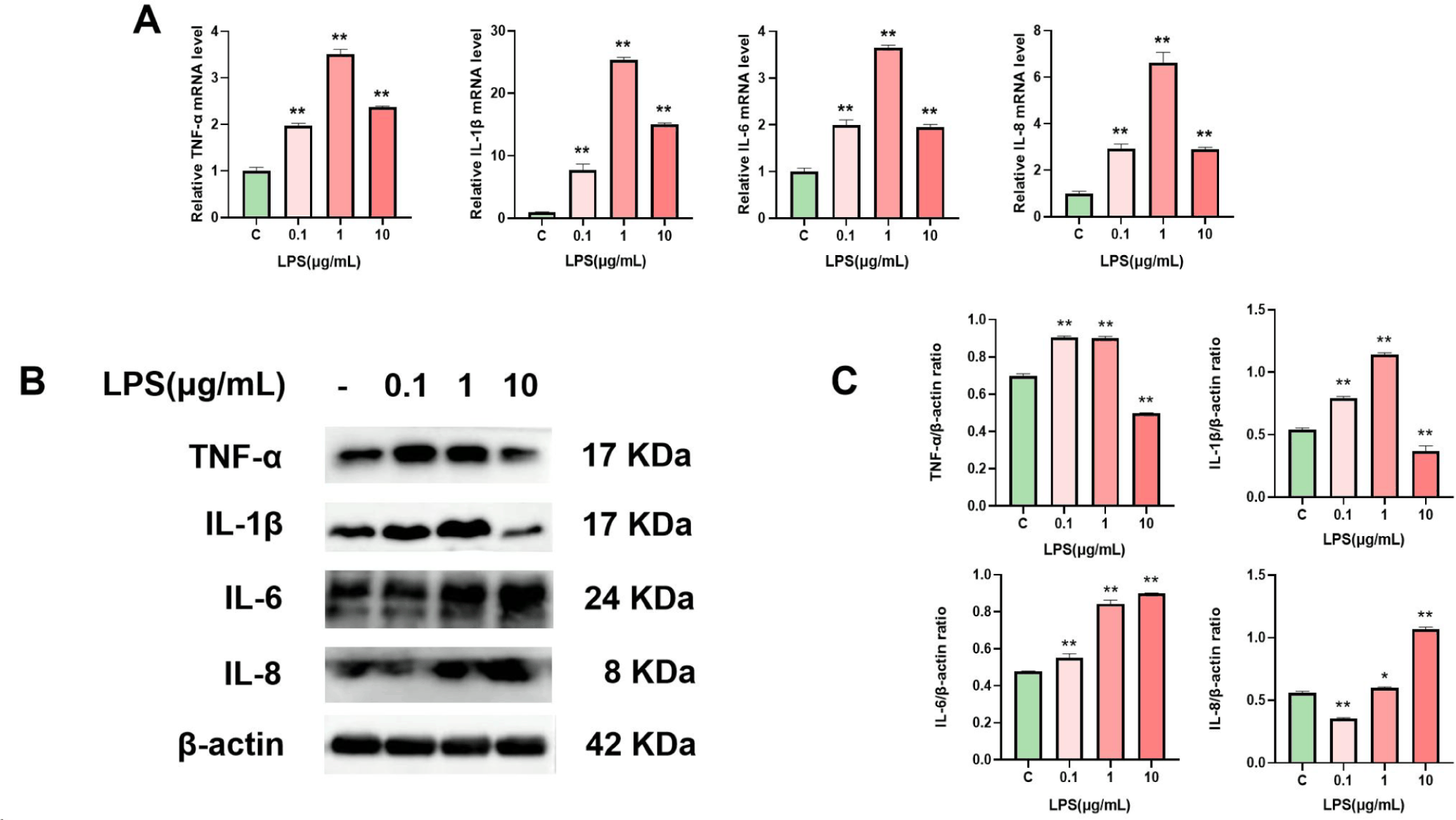
We used qRT‒PCR and WB to measure the mRNA and protein levels of TNF-α, IL-1β, IL-6, and IL-8 in LPS-treated cells to investigate the impact of LPS on the inflammatory response of BMECs. The findings demonstrated that at 1 µg/mL LPS, the mRNA (Fig. 2A) and protein (Figs. 2B and 2C) expression of cellular inflammatory components was considerably upregulated. Thus, the inflammatory model in this work was established by treating cells with 1 µg/mL LPS for 24 h.
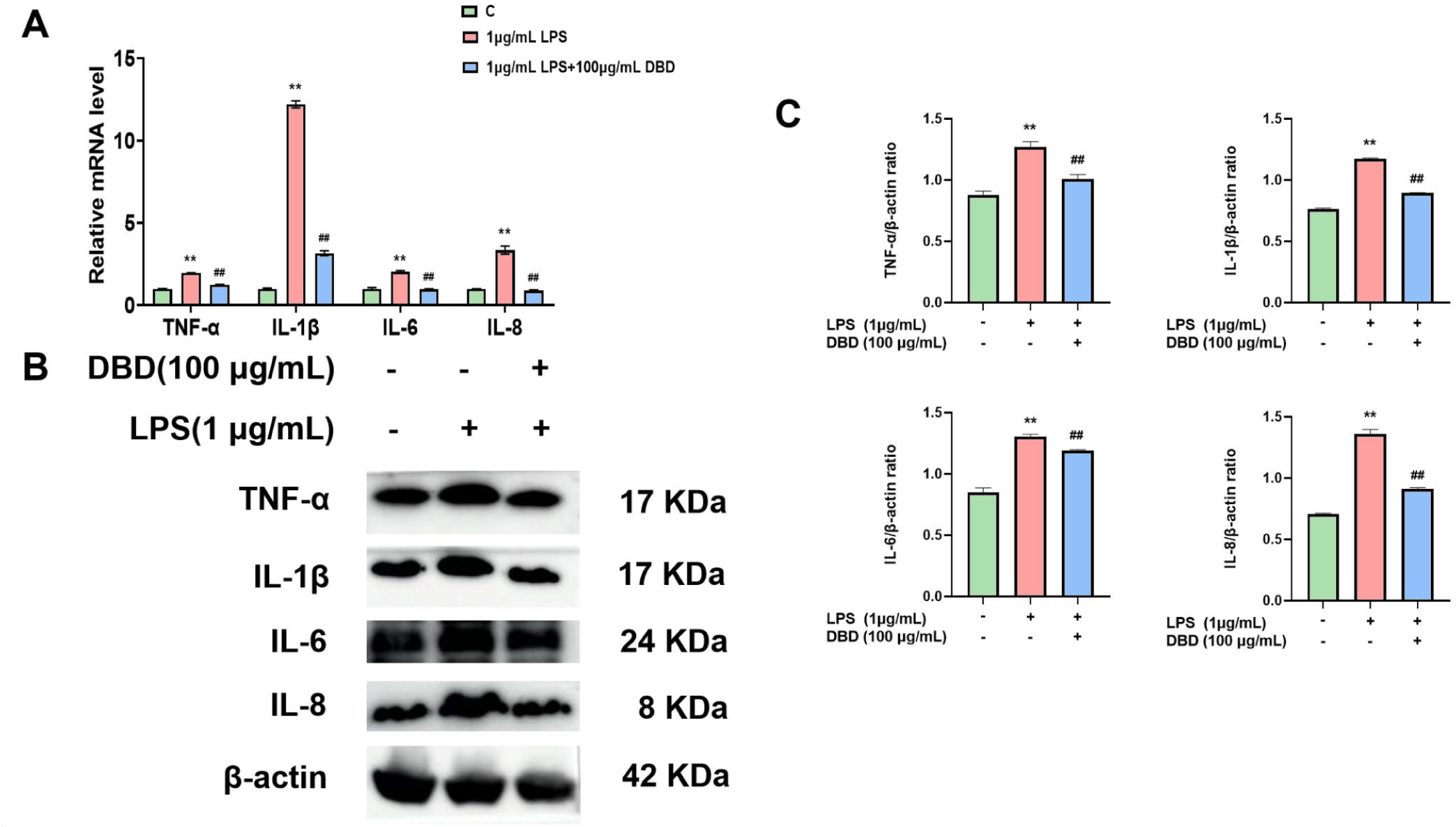
Using qRT‒PCR and WB, we investigated the mRNA (Fig. 3A) and protein (Figs. 3B and 3C) levels of TNF-α, IL-1β, IL-6, and IL-8 following LPS treatment to explore the anti-inflammatory effects of DBD on BMECs. Following LPS treatment, the expression levels of TNF-α, IL-1β, IL-6, and IL-8 were upregulated in comparison to those in the untreated group. On the other hand, the levels of these inflammatory factors decreased when DBD was present.
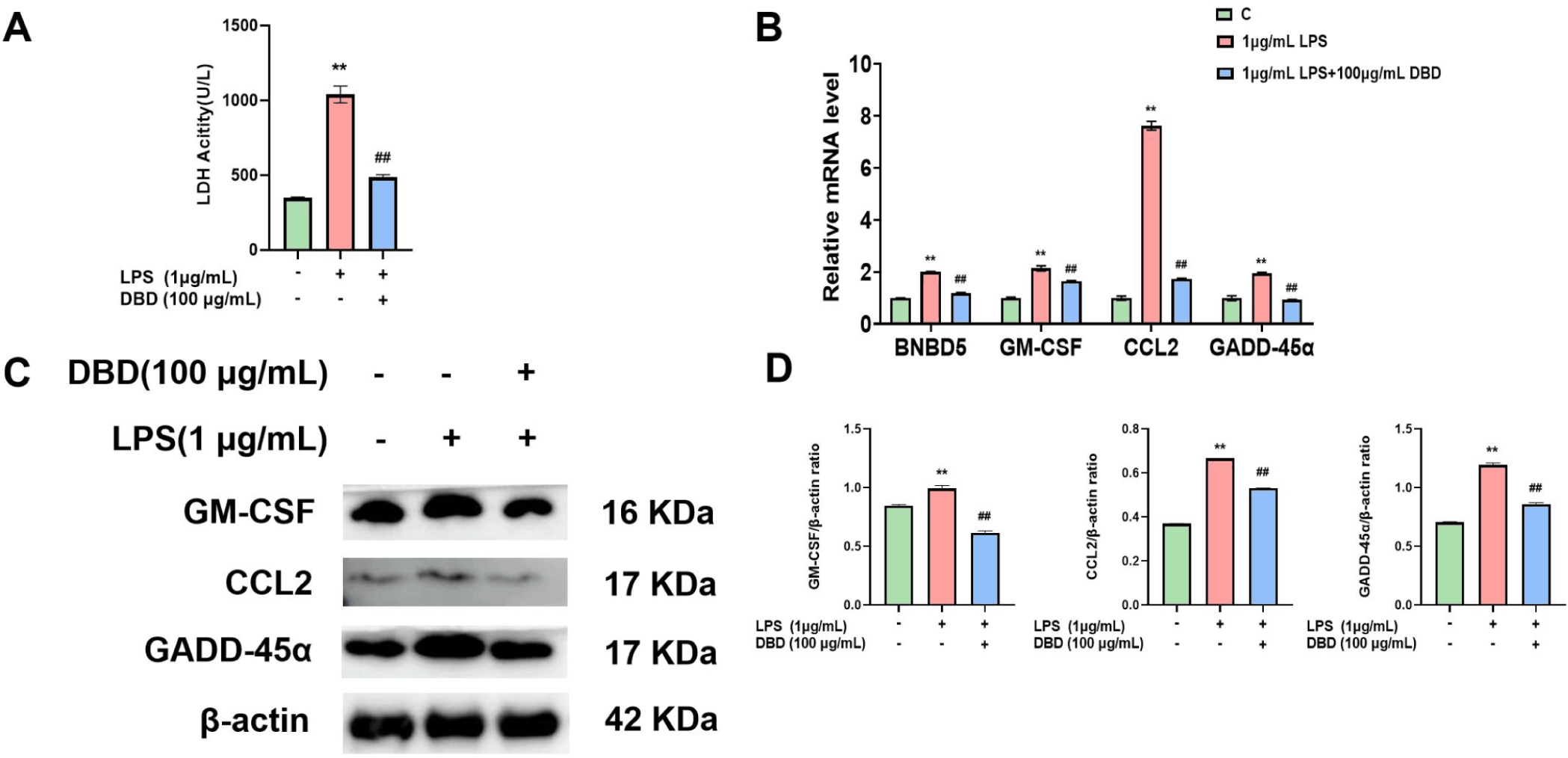
We observed the release of LDH within the fluid of LPS-treated cells, as well as the gene and protein expression of BNBD5, GM-CSF, CCL2, and GADD-45α within cells, to explore the protective impact of DBD against cell injury. LDH release was greater in the LPS group than in the control group. Similarly, the gene expression levels of BNBD5, GM-CSF, CCL2 and GADD-45α were elevated, and the levels of GM-CSF, CCL2, and GADD-45α were increased. However, in the presence of DBD, LDH release was reduced (Fig. 4A), and the gene (Fig. 4B) and protein (Figs. 4C and 4D) levels of cell-associated damage factors were decreased.
To investigate the ability of DBD to shield cells from oxidative damage, we detected the gene expression of COX-2, PPARγ, and iNOS in LPS-treated BMECs by DBD. At the same time, the contents of T-AOC, GSH, SOD and MDA in the cells were determined, the amount of NO in the cell culture supernatant was determined, and the reactive oxygen species staining was performed. The findings demonstrated that cells in the LPS group had increased COX-2, PPARγ, and iNOS gene expression; the content of T-AOC, GSH, and SOD in cells decreased significantly; while the content of MDA in cells increased, the concentration of NO in cell culture supernatant increased, and the level of reactive oxygen species increased. However, in the presence of DBD, the content of T-AOC, GSH and SOD in cells increased, while the content of MDA in cells decreased, the release of NO in cell culture medium decreased (Fig. 5A), the level of ROS in cells decreased (Fig. 5C), and the expression of oxidative stress-related factors decreased (Fig. 5B).
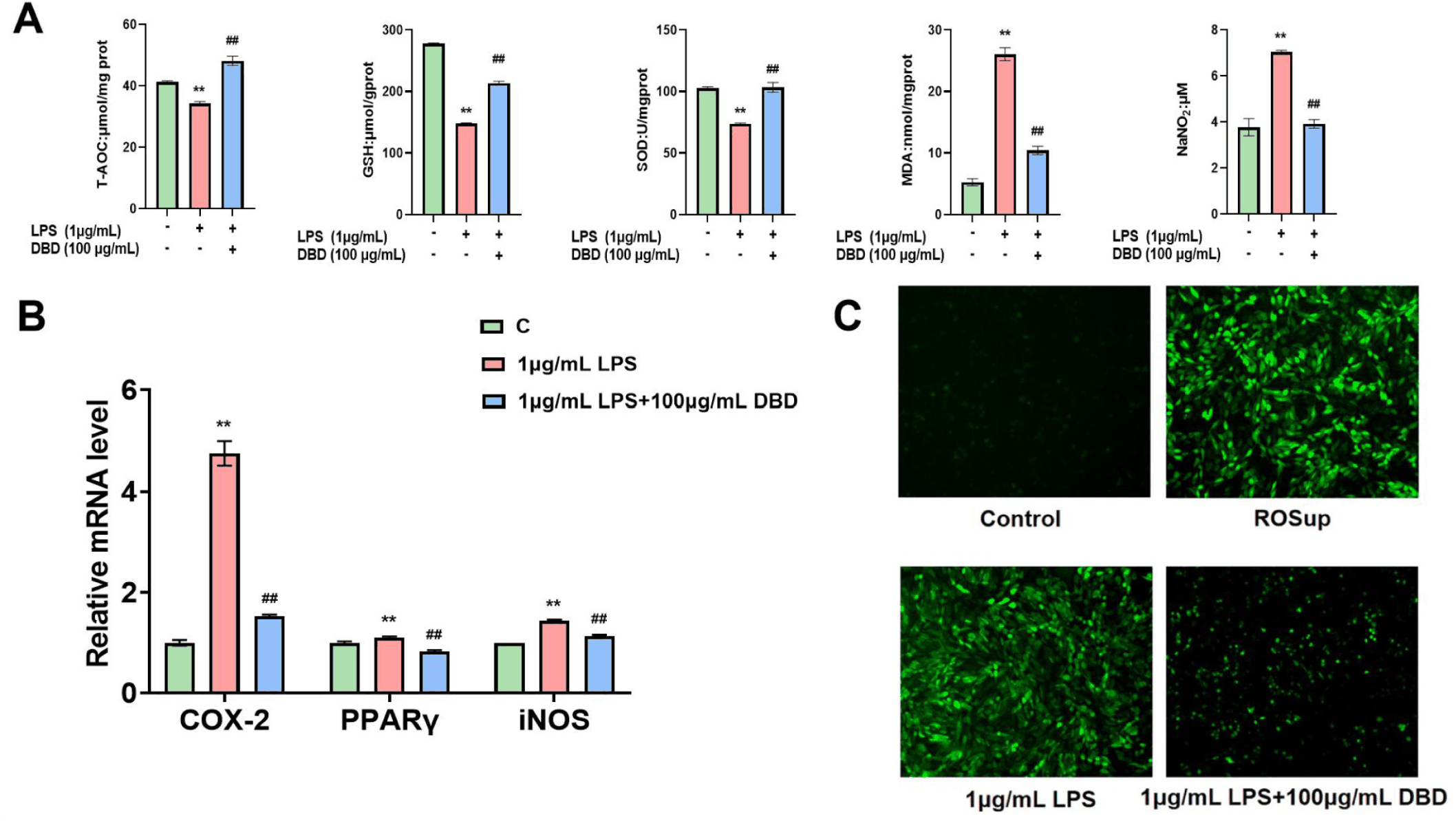
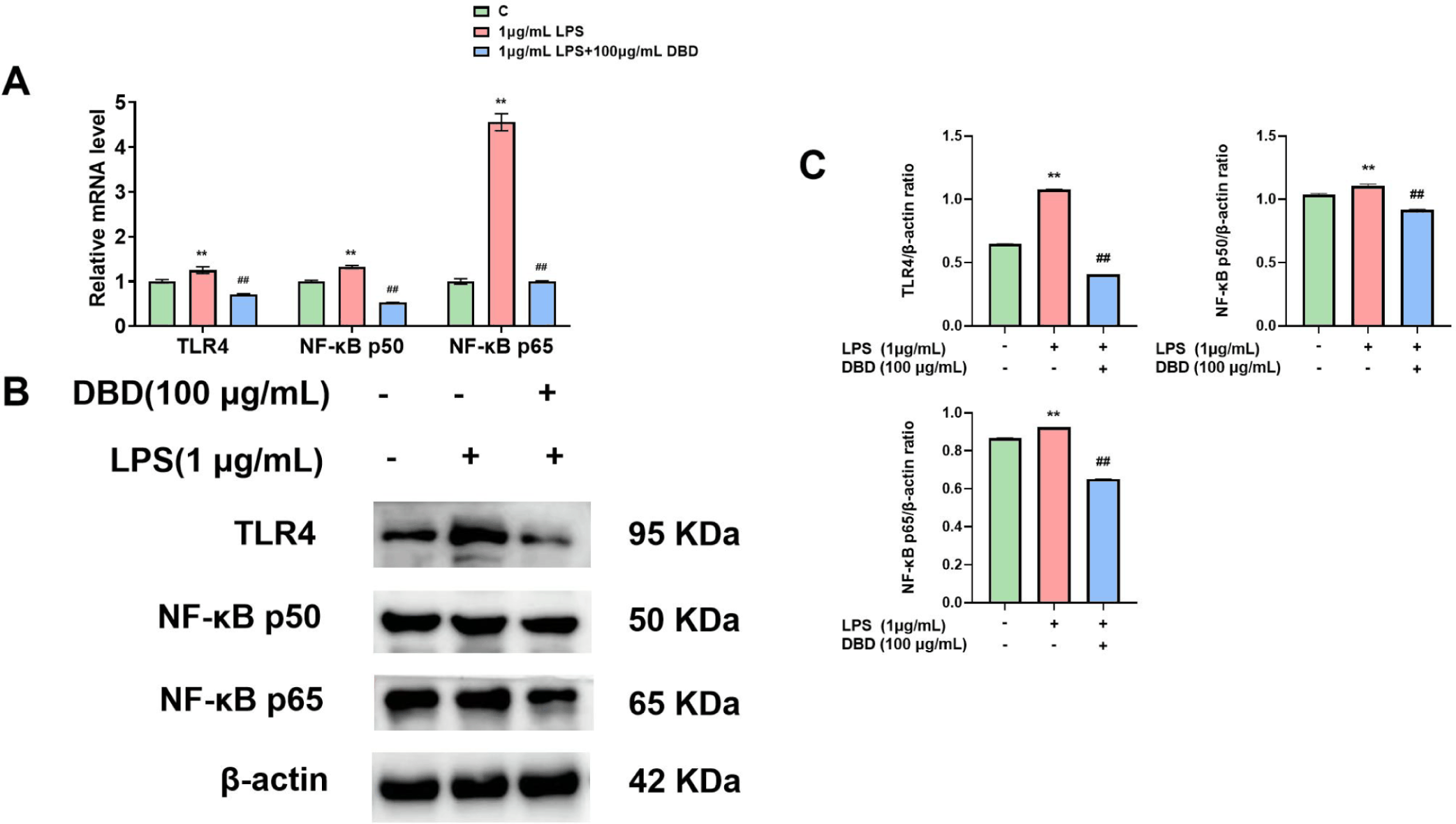
We evaluated the levels of NF-κB and TLR4 in vitro to determine the defense mechanisms of DBD on the mammary epithelial cells of inflamed cows. Cells in the LPS group exhibited increased TLR4 and NF-κB expression. On the other hand, TLR4 and NF-κB expression was downregulated when DBD was present (Figs. 6A and 6C).
DISCUSSION
Mastitis in dairy cows is a highly prevalent disease in the cattle industry that harms the wellbeing of cows, leading to decreased milk production, increased treatment costs, and compromised productivity [31]. In order to find a new alternative therapy for the treatment of BM, we used LPS to infect BMECs to establish an in vitro model of mastitis, and on this basis, we studied the protective effect of DBD on cells. Firstly, we observed the effects of 14 different concentrations of DBD on cell growth. The results showed that a low concentration of DBD could promote cell proliferation, while the high concentration of DBD could inhibit cell proliferation. The results of Choi et al. showed that DBD treatment could induce the proliferation of MG-93 cells and increase the number of cells compared with the treatment without DBD [32]. LI et al.’s study also showed that DBD could alleviate the decrease of cardiomyocyte viability induced by hydrogen peroxide and significantly increase cell viability [33]. Other studies have shown that DBD can also directly promote the growth of megakaryocytes [34], which is similar to our results. Therefore, we conclude that the active ingredients of traditional Chinese medicine in DBD can promote the absorption of nutrients by cells, and thus promote the proliferation of BMECs. The high concentration of DBD nutrients is too high, which will cause certain stimulation to the cells, cause the cell absorption burden, and thus inhibit cell proliferation.
Numerous investigations have demonstrated that mastitis is associated with an increase in the expression of intracellular inflammatory factors, including TNF-α, IL-1β, IL-6, and IL-8; therefore, mastitis is often used as an important indicator of the level of inflammation in dairy cows with mastitis [35–37]. In our study, we also found that LPS treatment upregulated the expression of TNF-α, IL-1β, IL-6, and IL-8 in BMECs. Several studies have confirmed the positive effects of herbal components on the control of mastitis in dairy cows. For example, Li et al. investigated the ability of matrine to prevent inflammation in BMECs and the underlying molecular mechanism, and the results showed that matrine pretreatment downregulated the expression of IL-1β, IL-6, IL-8, and TNF-α during inflammation in BMECs [38]. Song et al. investigated the effect of the ethanol extract of Artemisia annua (AAE) on LPS-induced inflammatory injury in BMECs and demonstrated that AAE treatment reduced the levels of several inflammatory factors in LPS-stimulated BMECs [39]. Another study revealed that LBP pretreatment significantly reduced the expression of TNF-α and IL-1β in cells stimulated with E. coli [40]. DBD, a traditional Chinese herbal formula, can exert potential immunoprotective effects by fighting inflammation, preventing injury and reducing oxidative stress and has been recognized as an effective immunomodulator [35,41,42]. Fang et al. reported that DBD suppressed skin inflammation and reduced the cellular levels of IL-4, IL-5, and TNF-α during the course of evaluating whether DBD could alleviate symptoms of atopic dermatitis [43]. Our study showed that DBD downregulated the expression of TNF-α, IL-1β, IL-6, and IL-8 after LPS-induced BMEC infection.
Cells experiencing injury exhibit increased LDH release and elevated expression of BNBD5, GM-CSF, CCL2, and GADD-45α [44–48]. Various studies have explored alterations in factors associated with injury in BMECs during inflammation and the impacts of pharmacological interventions. For example, BMECs are damaged by rupture of the cell membrane and release of LDH from the cytoplasm into the extracellular space, leading to increased LDH levels in cell cultures; however, in deoxynivalenol-induced inflammatory injury in mammary epithelial cells, treatment with taraxasterol strongly reduced LDH release [49]. In addition, BNBD5 is an important molecule in the innate defense system that activates the body’s first defense barrier and is considered a candidate gene for treating mastitis [50]. Studies have shown that BNBD5 is significantly upregulated in mastitis. GM-CSF acts as a “messenger” that promotes macrophage recruitment to the site of inflammation and increases the secretion of TNF-α and IL-6, directly inducing tissue injury [51]. CCL2 is a macrophage chemokine that functions by targeting immune cells for chemotaxis. Breast tissue injury occurs after S. aureus and E. coli infection, which leads to an intrinsic immune response in which the mRNA transcript level of CCL2 is upregulated [52]. Additionally, GADD-45α is a growth-blocking and DNA damage repair gene, and some studies have shown that LPS- or LTA-induced cellular inflammation in BMECs is accompanied by significantly increased expression of GADD-45α and DNA damage [53]. Therefore, researchers have noted that resistance to cellular damage is highly important for alleviating the development of inflammation. Using our LPS-induced mastitis epithelial cell model, we found that DBD reduced LDH release and downregulated the expression of the damage-associated factors BNBD5, GM-CSF, CCL2, and GADD-45α. These findings suggested that DBD can effectively ameliorate cellular injury and that this cellular injury is closely related to the generation of inflammation.
Recent investigations have established a strong relationship between oxidative stress levels and mastitis in dairy cows. Chen et al. highlighted the association between oxidative stress and bovine mastitis, noting that the excessive production of ROS by BMECs triggers apoptosis and the release of inflammatory factors. This suggests that a disruption in the intracellular redox balance impairs immune and anti-inflammatory functions in cows around the time of parturition [54]. In the context of the ability of betaine to mitigate inflammation and serve as an antioxidant, Zhao et al. reported that betaine decreased the LPS-induced inflammatory response and oxidative damage by reducing the expression of COX-2 and iNOS in LPS-stimulated BMECs [55]. Similarly, Fu et al. reported that Tanshinone IIA significantly lowered ROS and MDA levels in LPS-treated BMECs [56]. Additionally, Jiang et al. ‘s findings indicated that quercetin effectively shielded BMECs from LPS-induced oxidative and barrier damage [57]. Li et al. found that DBD could down-regulate the expression of IL-6 and TNF-α, reduce the level of MDA and increase the level of SOD in the process of using DBD to treat vascular aging in mouse with chronic intermittent hypoxia [58]. Our findings demonstrate that DBD, when applied to LPS-treated BMECs, it could increase the T-AOC of the cells, increase the GSH content, improve SOD activity, reduce the intracellular production of ROS, MDA, and NO, and down-regulate the expression of COX-2, PPARγ, and iNOS in order to protect the cells from oxidative stress damage. DBD has been used in China for more than 800 years. A large number of studies have shown that the main components of DBD are saponins, flavonoids, volatile oils, organic acids, polysaccharides and so on [13]. Ferulic acid is the main active component of DBD. Ferulic acid can not only reduce the formation of free radicals and ROS, increase the expression of antioxidant genes and the activity of antioxidant enzymes, but also promote the activity of G-CSF and accelerate the repair of damaged cells [26,59]. Total glucosides of DBD can improve pulmonary fibrosis and increase cell anti-fibrotic activity [60]. Quercetin in DBD belongs to flavonoids, which can inhibit the growth of cancer cells and prevent cancer [61]. Therefore, we believe that the anti-inflammatory, anti-injury and anti-oxidation effects of DBD are achieved with the help of ferulic acid, saponins, quercetin and other effective substances.
Extensive research has underscored the pivotal roles of TLR4 and NF-κB in managing mastitis [21,22]. LPS activation of NF-κB through the specific recognition of TLR4 suggested that NF-κB, an intracellular nuclear transcription factor, regulates the release of multiple inflammatory factors, inhibits cell proliferation, and promotes cellular inflammation. Consequently, TLR4 and NF-κB have been identified as potential therapeutic targets for treating mastitis in dairy cows [62]. Wang et al. reported that LPS stimulation of BMECs hindered cell proliferation, promoted the release of inflammatory and apoptotic factors, and increased the expression of TLR4 and NF-κB p65 [63]. Our experiments also revealed LPS-induced upregulation of TLR4 and NF-κB expression. Che et al.’s work with allicin showed that it could mitigate LPS-induced inflammation in BMECs by inhibiting the TLR4/NF-κB signaling pathway [64]. Moreover, β-carotene was found to reduce LPS-induced inflammation in BMECs by suppressing NF-κB expression [65]. In an investigation of artemisinin, it was observed that artemisinin blocked the TLR4-mediated NF-κB signaling pathway and decreased the expression of TNF-α, IL-6, and IL-1β mRNA in E. coli-induced BMECs, thus reversing the inflammatory response [66]. Our results indicated that DBD could downregulate the expression of TLR4, NF-κB p50, and NF-κB p65 following LPS induction in BMECs.
In conclusion, this study established an in vitro model of bovine mastitis by stimulating BMECs with LPS to explore the protective effects of DBD on these cells and achieved relatively satisfactory outcomes. DBD mitigated the LPS-induced expression of the inflammatory markers TNF-α, IL-1β, IL-6, and IL-8; reduced the expression of the damage indicators BNBD5, GM-CSF, CCL2, GADD-45α, and the release of LDH; and in terms of oxidative stress, DBD decreased the expression of COX-2, PPARγ, and iNOS, increased the T-AOC, GSH content, and SOD activity, lowered MDA content, and NO release, and attenuated the generation of ROS. These findings indicate that DBDs exhibit significant anti-inflammatory effects, provide protection against cellular injury, and alleviate oxidative stress. Additionally, DBD suppressed the expression of TLR4, NF-κB p50, and NF-κB p65, suggesting the potential of DBD as an effective treatment for mastitis in dairy cows. However, given the complexity of the immune response in animals, the in vitro model has certain limitations. Consequently, future research will involve validating the therapeutic efficacy of DBD through animal models of mastitis and clinical trials to investigate the mechanism by which DBD affects dairy mastitis more thoroughly.
CONCLUSION
LPS treatment of BMECs resulted in cellular inflammation, injury, and oxidative stress. DBD exerted anti-inflammatory, anti-injury, and antioxidative stress effects on LPS-treated BMECs. Furthermore, DBD significantly reduced the expression of TLR4 and NF-κB in these cells. These findings suggest the potential of DBD as an innovative treatment for mastitis in dairy cows. Consequently, DBD has been proposed as a promising new therapeutic option for managing dairy cow mastitis based on these findings.