INTRODUCTION
Soybean (Glycine max), an annual crop belonging to the Leguminosae or Fabaceae family, is the most economically important bean globally. Its essential components such as a relatively high digestible protein, dietary fibre, free sugar, minerals, and essential fatty acid composition [1], and a high and well-proportioned amino acid (AA) profile, with the exception of sulfur-containing amino acids [2], are what make it a widely accepted feedstuff. It is primarily used to produce oilseed meal or vegetable oil for livestock feeding [3,4]. Soybean meal (SBM) is the residue left over after soybean oil is extracted mechanically and solvently, contains about 46%–48% crude protein, 2.5%–3.5% lysine, 0.6%–0.7% tryptophan, 0.5%–0.7% methionine, and 0.5%–0.8% cystine [5]. It may be without of unpredictable antinutrients if processed properly, is accessible year-round, has little change in nutrient composition, provides a limited amount of animal protein, such as fish and blood meal, and is often chosen when creating diets. Furthermore, several anti-nutritional factors found in SBM, including oligosaccharides, trypsin inhibitors, and antigenic proteins, interfere with nutrient absorption and digestion, negatively impacting the growth performance, feed utilization, and physiological status of pigs [6], while also resulting in oxidative stress and inflammation linked to compromised gut function [7]. According to Muniyappan et al., microbial fermentation raises the nutritional quality of SBM by decreasing the ANF content and boosting nutrient bioavailability [8]. The negative physiological reactions of the gastrointestinal systems are mostly caused by interacting antigenic proteins and several soybean anti-nutrients with the gastrointestinal tract [9]. Microbial fermentation is a processing method for soybean meal that can remove certain macromolecular nutrients, non-digested sections, and antinutritional substances. Fermented soybean meal (FSBM) is made from SBM using yeast, fungal species such as Saccharomyces cerevisiae and bacterial species, such as Lactobacillus sp., Bacillus sp., Aspergillus sp., and Streptococcus sp. [10–12]. Previous studies have shown that the primary antigenic protein in soybeans, β-conglycinin and glycinin, may be broken down into smaller peptides with antioxidant potential FSBM [13,14]. FSBM production from microorganisms is a unique and effective feed method that could reduce antinutritional factors, break down significant nutrient molecules, create bioactive chemicals, and alter the gastrointestinal tract to ultimately increase digestibility [11]. Dietary supplementation of FSBM could increase growth performance, nutrient digestibility, and feed efficiency [15]. This review summarizes the current status of swine nutrition and describes how the use of fermented feed can improve growth performance, nutrient utilization, gut health, and immune responses in swine diets.
FERMENTED SOYBEAN MEAL PROCESSING
Fermentation, which is traditionally used to preserve or improve feed quality, has recently gained attention within the livestock industry due to its effectiveness in feed processing. This method effectively eliminates toxins and antinutrients while breaking down macromolecules through the action of microbes, resulting in the formation of bioactive compounds and metabolites. Temperature, pH, nature and composition of the medium, dissolved O2 and CO2, operating systems (e.g., batch, fed-batch, continuous), addition of precursors, mixing (cycling in different environments), and fermentation shear rates and length of the fermentation process can affect the fermentation rate and the quality of the fermentation products [8]. Depending on the type of microorganisms involved, fermentation will produce different end products such as lactic acid, ethanol or acetic acid, as different microorganisms may react differently to each substrate [15]. Lactobacillus produces lactic acid, mold produces citric acid, whereas yeasts produce ethanol and CO2 [11]. It serves as a metabolic mechanism that improves the absorption of minerals and converts sugar into energy. These complexes are usually broken down by enzymes that require fermentation to maintain optimal pH [16]. Following the European Union’s ban on antibiotics as antimicrobial growth promoters in livestock, interest in feeding fermented feeds to improve animal health has increased dramatically [17]. During the fermentation process of SBM, Lactobacillus plantarum, Enterococcus faecalis, Bifidobacterium bifidum, Lactobacillus acidophilus, Bacillus subtilis, S. cerevisiae, Bacillus licheniformis, Rhizopus oligosporus, Aspergillus oryzae, Enterococcus faecium, and Neurospora crassa play a crucial role [18,19]. According to a previous study, fermentation of SBM with L. plantarum, Enterococcus faecium, Aspergillus oryzae, and B. subtilis significantly reduced antinutrients such as phytate, trypsin inhibitors, and protease inhibitors [5]. Additionally, studies conducted by Zhang et al. found that fermentation of SBM with B.subtilis and lactic acid bacteria favored the production of antimicrobial peptides while facilitating lactic acid production, which improves feed acidity and enhances palatability [20]. Microbial Enzymes and Corresponding ANF Targets (Table 1).
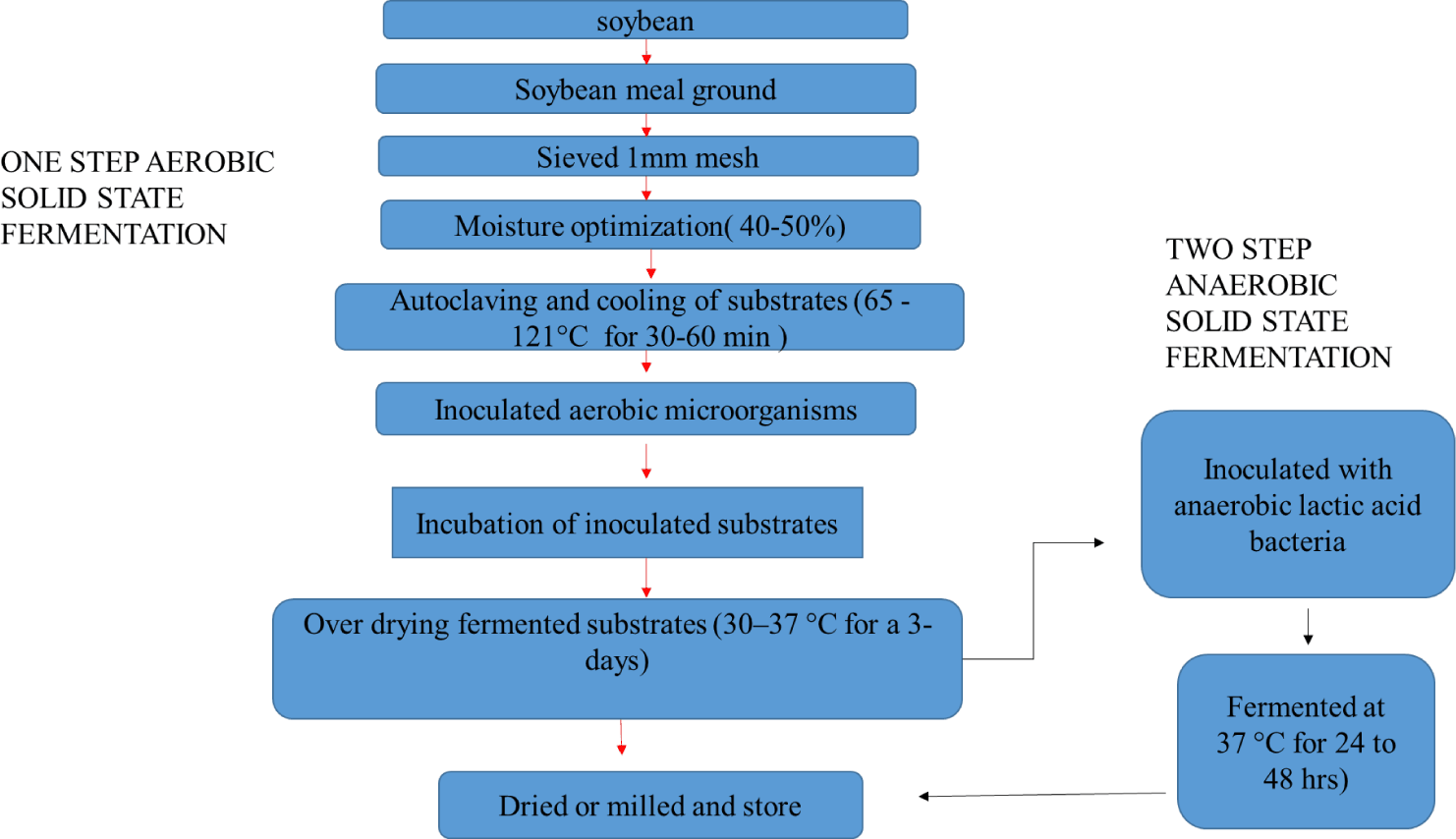
There are two main reasons why fermentation is necessary for soybeans. First, during fermentation, probiotic bacteria proliferate and produce metabolites with bioactive properties by utilizing non-protein anti-nutritional factors such as phytic acid, oligosaccharide, non-starch polysaccharides, and thyroxine [21]. Additionally, when fed, these probiotics transport microbes into the animal’s gut, where they modify the gastrointestinal microbial community to improve digestion [22]. Alternatively, a more prominent area in most research is that microbes release certain proteases to break down the anti-nutritional protein in SBM. Fermented feeds can be produced through different methods, including ensiling, liquid fermentation, and solid-state fermentation, which have gained popularity in recent years [23]. Solid-state fermentation, a conventional technique with a long history in food production and multi-organism work, is an integral part of FSBM processing. This method utilizes low humidity levels, which reduces the drying time of the protein hydrolysates. Studies have shown that it produces more metabolites than submerged fermentation, including digestive enzymes and high-value bioproducts such as bacterial antimicrobial peptides and antibiotics. Moreover, solid-state fermentation offers the advantages of relatively small amounts of pre-treatment and a wider culture environment. Due to low capital requirements and environmental friendliness, large-scale production is possible [24,25]. Additionally, solid-state fermentation can be a one-step process with a single fermentation phase or a two-step process with two-phase fermentation [26].
Although several studies have investigated the use of microbial additions to facilitate one-step direct fermentation of soybean [27], two-step solid fermentation is more successful. This method involves anaerobic fermentation during the second step and aerobic fermentation in the first (Fig. 1 and Table 2). Various bioactive compounds, including vitamins and enzymes, are produced by aerobic microorganisms such as Bacillus, Lactobacillus and fungi during the initial stages of aerobic fermentation, which consecutively stimulates the growth of lactic acid bacteria. According to Qiu et al., in practice, SBM is coarsely crushed, and sieved through a 1-mm mesh, followed by the addition of sterile water to achieve a moisture content of 37% [19]. Aerobic bacteria such as Lactobacillus sp. and Bacillus sp. are then inoculated, and the SBM undergoes fermentation at 32°C–37°C for four days. This is followed by anaerobic solid-phase fermentation resulting in the production of more lactic acid and promoting the growth of lactic acid bacteria. Under anaerobic conditions, microorganisms are forced to lyse, leading to the release of intracellular enzymes and other biological components. In this case, the already aerobically fermented SBM is re-inoculated with lactic acid-degrading bacteria, sealed in plastic bags, and allowed to undergo anaerobic fermentation at 32°C to 37°C for two to three days, after which it is dried, milled and stored. Additionally, Muniyappan et al. determined the effectiveness of solid-state fermentation in enhancing the nutritional quality of SBM and reducing its anti-nutrient content [8].
| Microbes | Inoculant dose | Moisture content (%) | Duration of fermentation | Reference |
|---|---|---|---|---|
| Bacillus subtilis BS12 | 108 CFU/mL | 50 | 37°C for 24 h | Zhang et al. [28] |
| Enterococcus faecium SLB120 | 1.0 × 108 CFU/g | 10 | 37°C for 36 h | Jeong et al. [11], Muniyappan et al. [18] |
| Lactobacillus plantarum, Bacillus subtilis, and Saccharomyces cerevisiae | 108 CFU/g | 10 | 37°C for 24 h | Zhu et al. [12] |
| Aspergillus oryzae GB-107 | 108 CFU/g | 30–40 | 37°C for 48h | Cho et al. [29] |
| Aspergillus oryzae 3.042 | 10,000 CFU/g | 10 | 80°C for 3 days | Feng et al. [10] |
| Bacillus subtilis CP-9 | NS | 30–40 | 37°C for 3 days | Akhtar et al. [30] |
| Bacillus subtilis, Lactobacillus, and Saccharomyces cerevisiae | NS | 10 | 30°C–37°C for a 3-days | Yan et al. [31] |
| Streptococcus thermophilus (CGMCC No. 1.2471), Saccharomyces cerevisae (CGMCC No. 2.1793), and Bacillus subtilis MA139 | 1 × 107 CFU/mL | 30–40 | 37°C for 24 h | Wang et al. [32] |
| Aspergillus oryzae and Lactobacillus reuteri | NS | 10 | 37°C for 24 h | Luo et al. [33] |
| Bacillussubtilis QB8 and Lactobacillusplantarum QP28-1 | NS | 30–40 | 30°C–35°C for a 5-days | Zhang et al. [20] |
| Saccharomyces cerevisiae, Bacillus subtilis, Bacillus licheniformis, Lactobacillus plantarum, and Lactobacillus reuteri | 4 × 107 CFU/mL and 2 × 107 CFU/mL | 37 | 32°C–37°C for 4 days | Qiu et al. [19] |
GROWTH PERFORMANCE
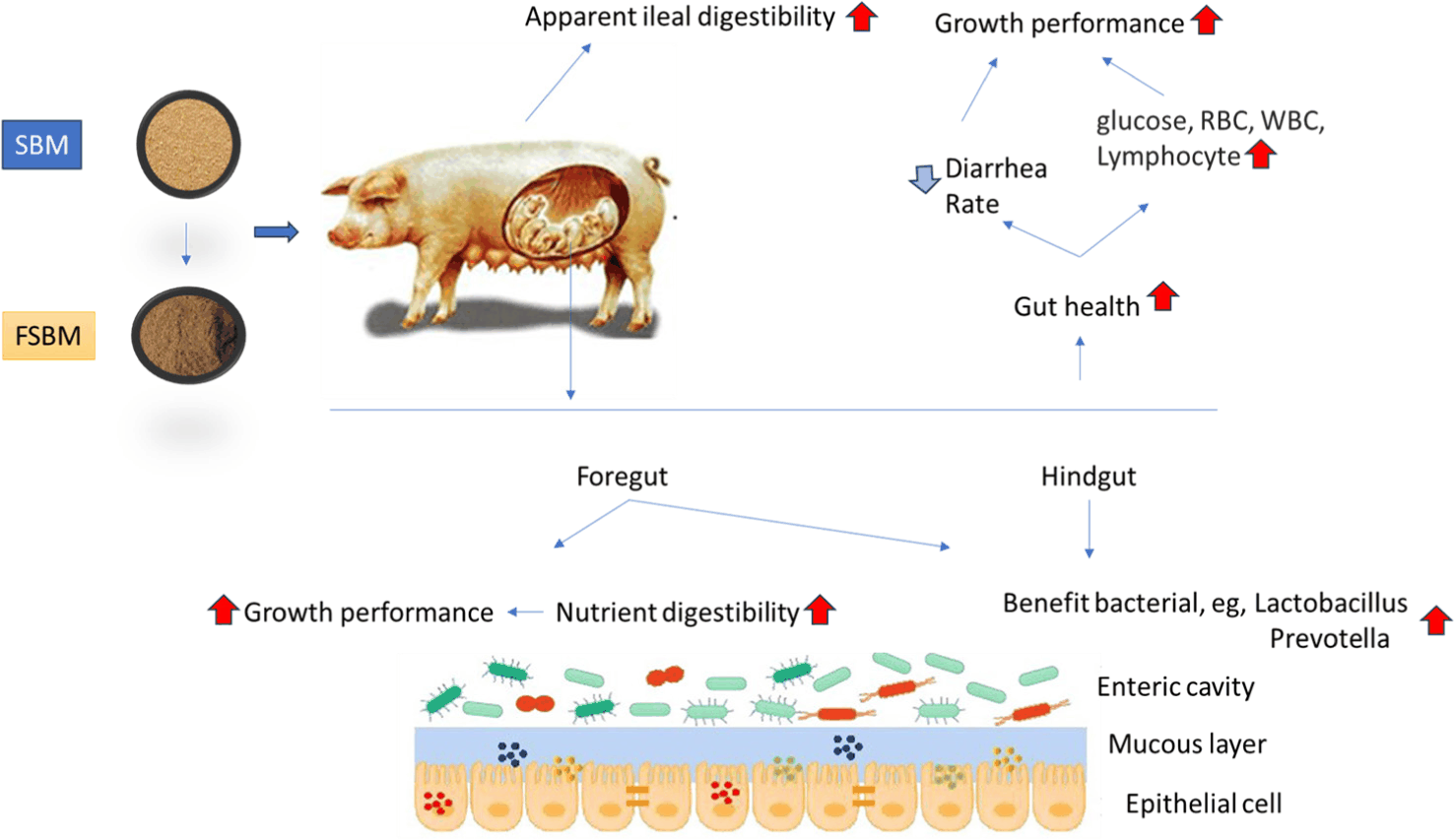
The dietary application of various microorganisms and their fermented soybean meal have been studied as a potential tool to improve growth performance and reduce mortality in pigs [34]. The mechanism by which FSBM may boost swine growth, includes the increase of endogenous digestive enzymes, saliva, bile, and mucus, lowering the abundance of harmful bacteria in the GIT, or modifying intestinal morphology through anti-inflammatory and antioxidant activities [35]. Furthermore, FSBM derived from bacteria, fungi, and yeast can potentially decrease antinutritional components in feed, equally boosting feed intake and growth performance [27]. Feeding weaning piglets soybean meal fermented with Enterococcus faecium has been shown to increase growth performance [11,18]. However, in another study, no health effects on piglet performance were observed by Lan and Kim, when SBM was replaced with up to 5% SBM fermented with Enterococcus faecium [36]. Soybean meal fermented with Streptococcus thermophilus, B. subtilis, and S. cerevisiae could increase (p < 0.05) the average daily gain (ADG) and average daily feed intake (ADFI) in weanling piglets [37]. In an experiment assessing the effects of SBM fermented with B. subtilis in pigs, Feng et al. observed that growth performance was greater than unfermented SBM [38]. Similarly, increased (p < 0.05) body weight gain, ADG (0.5%) and ADFI and decreased gain feed ratio (G:F) were recorded in pigs fed with SBM fermented with B. subtilis [39]. Piglets fed FSBM showed significant improvements (p < 0.05) in ADG (0.7%), lower G:F, elevated serum alkaline phosphatase and total serum protein levels, decreased serum urea nitrogen, and improved intestinal morphology. This was evidenced by reduced crypt depth, and enhanced villus height in the duodenum, jejunum, and ileum, as well as an increase in concentrations of serum immunoglobulins IgG, IgM, and IgA [12]. Compared with the SBM, pigs fed with FSBM showed higher (p < 0.05) body weight and ADG (2.03%) [13]. Gebru et al. reported that dietary supplementation of FSBM could improve (p < 0.05) ADG (0.33%) and G:F and lower serum haptoglobin in growing pigs [40]. Additionally, dietary supplementation of FSBM was shown to work synergistically to increase energy uptake and hepatic energy retention, the change in energy may occur due to increased (p < 0.05) ADG (2%) and G:F in nursery pigs [41]. Xu et al. reported an increased (p < 0.05) body weight gain and decreased G:F in pigs compared to those fed SBM [42]. The dietary inclusion of SBM fermented with L. plantarum, B. subtilis, and S. cerevisiae showed higher (p < 0.05) final body weight, ADG (44%) and ADFI, accompanied by a lower G:F in weaned piglets [43]. FSBM increased the generation of short-chain fatty acids and produced immunomodulation due to changes in microbial populations in the gastrointestinal tract, which also improved energy metabolism [43]. The metabolites of SCFA, as developed by bacteria digest carbohydrates in the small intestine, and affect leukocytes and endothelial cells by inhibiting histone deacetylase and activating G-protein-coupled receptors. In addition to their interactions with different receptors, SCFAs suppress the NF-κB transcription factor, decrease the synthesis of chemokines and cytokines, and encourage B-immune cells to produce IgA. Rojas and Stein. demonstrated that the dietary inclusion of FSBM can increase (p < 0.05) feed efficiency and growth performance in pigs [44]. Additionally, Zhang et al. reported an improved (p < 0.05) ADG (0.8%) and ADFI and lower G:F in piglets fed with SBM fermented with L. plantarum, B. subtilis compared to SBM [20]. Dietary inclusion of FSBM increases (p < 0.05) ADG (0.5%), immune function and gut microbiota in piglets [45]. Canibe and Jensen. reported an increase (p < 0.05) in body weight, ADG and ADFI in pigs fed diet supplemented with FSBM compared to SBM [46]. Wang et al. [32] also showed that FSBM inclusion in the pigs significantly improve (p < 0.05) ADG (34%) and decreased G:F and diarrhea compared to SBM when challenged with Enterotoxigenic Escherichia coli [32,47]. A 14 days experiment showed that dietary supplementation of FSBM could increase (p < 0.05) ADFI and decrease serum cortisol levels of piglets challenged with Lipopolysaccharide [48]. Canibe et al. and Kim et al. [49,50] reported that FSBM supplementation improved (p < 0.05) growth performance in weaned pigs. The results indicated positive effects on the intestinal T-cell immune system. Similarly, FSBM produced differences in feed intake and body weight, though it produced expected histomorphological changes in the gut. Lastly, in comparison with SBM, the dietary inclusion of FSBM resulted in increased (p < 0.05) growth performance in weaned pigs [51]. Complex proteins and carbohydrates are broken down through fermentation into smaller, more absorbable nutrients. Consequently, animals can extract more nutrients from the same quantity of feed. Antinutrients present in raw soybean meal, such as phytic acid and trypsin inhibitors, prevent digestion. Fermentation improves the function of the digestive tract by eliminating or reducing these antinutritional factors. Beneficial bacteria and organic acids that enhance gut health are often found in fermented feeds. A healthy gut promotes better development by preventing illness and improving nutrient absorption. Amino acids serve as the building blocks of growth, and fermentation makes them more accessible. This indicates that the body can utilise more protein to build muscle and tissue.
In contrast, dietary supplementation of FSBM could not increase growth performance and protein bioavailability in weaning piglets which can be as a result of the varying supplementation levels and form of inclusion in the diets of piglets [41]. Cheng et al. observed no significant effect (p < 0.05) with ADG, ADFI, and G:F in piglets fed with SBM fermented with L. acidophilus, Lactobacillus delbrueckii, Lactobacillus salivarius, and Clostridium butyricum compared to SBM [22]Click or tap here to enter text.. Additionally, When pigs were used to investigate the effects of fermented with L. plantarum, and B. subtilis on the gut microbial profile and growth performance, FSBM supplementation did not significantly improve (p < 0.05) BWG, ADFI, and G:F [51]. The different results could be due to differences in diet composition, animal age, and animal status.
NUTRIENT DIGESTIBILITY
Utilization of nutrients may vary on the components of FSBM present in the supplied feed as well as the functionof the gut. The composition of the FSBM portion of a feedstuff will determine how it behaves once ingested. The FSBM solubility and water holding capacity determine its viscosity, and fermentability impacts lower gut utilization and health. High viscosity will decrease the rate of endogenous enzyme diffusion into the digesta, which will reduce nutrient digestion. Additionally, highly viscous digesta will have less interaction with the brush border membrane enzymes, which also decreases digestibility and nutrient utilization. Feng et al. reported that SBM fermented by Aspergillus oryzae could increase the digestibility of dry matter (DM; 97.41% vs 88.17%), and crude protein (CP; 49.41% vs 43.54%) compared to SBM [10]. Further studies indicated that SBM fermented with Enterococcus faecium showed improved DM (84.52% vs 81.10%), CP (84.45% vs 81.96%), and gross energy (GE; 82.72% vs 80.78%) digestibility in weaning pigs [18]. Furthermore, Muniyappan et al. found that dietary supplementation of SBM with Enterococcus faecium resulted in a significant increase in apparent total tract digestibility of DM (83.40% vs 80.43%), nitrogen (84.43% vs 81.85), and GE (82.69% vs 80.85%) and apparent ileal digestibility DM (74.91% vs 74.54%), nitrogen (78.28% vs 75.26%) and GE (76.48% vs 74.99) in weaning pigs [8]. Moreover, Jeong et al. showed that dietary supplementation of SBM with Enterococcus faecium resulted in a significant increase in apparent ileal digestibility of DM (85.0% vs 82.9%), nitrogen (83.2% vs 78.3%) and GE (84.6% vs 78.0%) in weaning pigs [11]. Ma et al. reported that dietary inclusion of FSBM can increase CP (73.1% vs 75.0%) and amino acid levels while lowering the presence of trypsin inhibitor, β-conglycinin, and glycinin compared to SBM [52]. Additionally, Rojas and Stein. reported that the dietary supplementation of SBM fermented by Aspergillus oryzae and B. subtilis results in increased digestible energy, metabolizable energy, net energy and digestibility of GE (88.2% vs 84.0%) and nitrogen (91.3% vs 76.0%) and apparent ileal digestibility of CP (65% vs 60%) in weaning pigs [44]. Yuan et al. also reported improvements in CP (87.44% vs77.43%), crude fat (71.08% vs 48.45%), calcium (82.2% vs 69.89%) and phosphorus (65.75% vs 57.76%) of piglets fed dietary supplements of SBM fermented with Lactobacillus casei, B. subtilis, and Hansenula anomala [15]. Furthermore, Wang et al. indicated that the dietary inclusion of Streptococcus thermophiles, S. cerevisiae, and B. subtilis fermented SBM in weaning pigs enhances the digestibility of GE (80.88% vs 83.40%) and the ileal digestibility of CP (70.10% vs 73.87%) [37]. In SBM, intact cell wall matrices mainly cellulose, hemicellulose, and pectin physically enclose nutrients (such as proteins and carbohydrates). During digestion, this structural barrier restricts enzymatic access, which lowers the bioavailability of nutrients. Encapsulated proteins and lipids are released when fermentation breaks down the cell wall. Pigs’ crude protein digestibility is between 15 and 25 percent (compared to non-fermented soy). Antinutritional components (such phytic acid) are simultaneously exposed to microbial phytases due to cell wall disintegration, which accelerates their decomposition. Compared with the SBM, the dietary supplementation of FSBM also improved DM (82.14% vs 84.33%), GE (82.05% vs 84.30), CP (73.75% vs. 80.84%), ether extract (63.09% vs 67.35%), ash (54.18% vs 59.14%) and calcium (56.93% vs 58.52%) of weaned pigs [50]. Moreover, Akhtar et al. observed that the dietary inclusion of SBM fermented with B. subtilis increases the ileal digestibility of CP (84.2% vs. 78.3%), ash (55.6% vs. 36.1%), DM (80.0% vs. 71.7%), ADF (62.4 vs. 53.3%), and NDF (70.9% vs. 66.0%) and apparent total tract digestibility of DM (89.5 vs. 87.2%), CP (86.3 vs. 83.7%), GE (88.0 vs. 87.5%), (48.3 vs. 36.1%), NDF (86.0 vs. 78.5%), and ADF (79.8 vs. 73.0%) in growing pigs [30]. It has been shown that the inclusion of FSBM could increase DM (86.43% vs. 84.19%), GE (85.74% vs 83.66%), CP (77.81% vs 71.09%), calcium (61.30% vs 54.34%), ether extracts (61.30% vs 53.05%) and phosphorus (58.18% vs 50.89%) compared to SBM [51]. Upadhaya and Kim. noted improved apparent ileal digestibility of nitrogen (77.2% vs. 74.3%) and standardized ileal digestibility of CP (87.79% vs.84.81%) in weaning pigs fed diets supplemented with SBM fermented by B. subtillus, as well as SBM fermented with Saccharomyces carlsbergensis and SBM fermented with Saccharomyces carlsbergensis and Bacillus amyloliquefaciens compared to SBM [53]. Hossain et al. reported that the dietary inclusion of B. subtilis fermented SBM in weanling pigs leads to improved apparent total tract digestibility and ileal digestibility of DM (82.40% vs. 78.45%), nitrogen (82.54% vs.79.33%), and GE (81.26% vs.76.77%) and standardised ileal digestibility of CP (88.11% vs. 84.32%) compared with the SBM [54]. The dietary inclusion of B. subtilis fermented SBM in pigs could increase DM, CP, GE, ether extracts and ash digestibility [12].
In contract, Cho et al. reported that the dietary inclusion of SBM fermented with Aspergillus Oryzae did not affect the digestibility of DM and nitrogen compared to SBM [29]. No difference was observed in the nutrient digestibility of weaning pigs fed an FSBM-supplemented diet [55]. Wang et al. observed no effects on nutrient digestibility in weaning pigs on diet supplemented by SBM fermented with Streptococcus thermophiles, S. cerevisiae, and B. subtilis [37]. The dietary supplementation of SBM fermented with Aspergillus Oryzae and B. subtilis had no influence on DM, CP, energy, and calcium digestibility in weaned pigs [56].
Possible mechanisms resulting in improved nutrient digestion by FSBM supplementation include the ability of these nutrients to stimulate appetite, salivary secretion, bile acid secretion, intestinal mucus production, and the activity of digestive enzymes such as amylase and trypsin and positively affect the gut morphology [12]. Alternatively, FSBM may act in an overlapping mode of action involving local effects at the gut boundary and systemic alterations of macronutrient metabolism [23].
AMINO ACID DIGESTIBILITY
Nutrient utilisation may differ depending on the gut’s function and the DF components in the diet. The amount of β-mannan in DF found in SBM is comparatively high [27]. The response of a feedstuff after ingestion is determined by the composition of its NSP component. The viscosity of the fibre is determined by its solubility and water-holding capacity, whereas fermentability affects gut health and utilization. Reduced nutrient digestion will result from high viscosity because it slows the pace at which endogenous enzymes diffuse into the digesta. Furthermore, excessively viscous digesta will interact less with the enzymes in the brush border membrane, reducing digestibility and nutrient utilisation. SBM contains 233 g/kg of dietary fermentation with microorganisms enhance utilization within the gastrointestinal tract. Several studies have explored the effect of FSBM on amino acid digestibility, which helps to recognize nutrient metabolism and growth performance [18,56]. Jeong et al. reported a significant increase in ileal amino acid digestibility, including arginine, lysine, aspartic acid, glutamic acid, glycine and tyrosine due to the dietary supplementation of SBM fermented by Enterococcus faecium [11]. Furthermore, Muniyappan et al. reported that dietary inclusion of Enterococcus faecium fermented SBM in weaning pigs led to an increase in apparent total tract digestibility of valine, isoleucine, phenylalanine, arginine, glutamic acid, proline, total non-essential amino acids and total amino acid as well as the ileal digestibility of valine, isoleucine, phenylalanine, lysine, arginine, tryptophan, serine, glutamic acid, proline, glycine, cysteine [8]. Cervantes-Pahm and Stein. indicated that supplementation with FSBM showed an improvement in the apparent ileal digestibility of histidine, iloleucine, lysine and phenylalanine and standardized ileal digestibility of lysine and phenylalanine in weaned pigs [57]. Similarly, Yun et al. reported an increase in apparent ileal digestibility of histidine, iloleucine lysine lysine, phenylalanine, valine, glycine, serine, and proline, as well as improvements in apparent fecal amino acid digestibility of arginine, histidine, isoleucine, lysine, methionine, phenylalanine, threonine, alanine, cystine and serine in weaned pigs fed diets supplemented with FSBM [51]. Upadhaya and Kim. observed improvements in apparent and standardized ileal amino acid digestibility of lysine, phenylalanine, asparatic acid, glutamic acid, glysine, and serine in diets supplemented with SBM fermented with B. subtillus, Saccharomyces carlsbergensis, and a combination of Saccharomyces carlsbergensis and B. amyloliquefaciens compared to unfermented SBM [53]. Hossain et al. reported that dietary inclusion of SBM fermented with B. subtilis had beneficial effects on apparent total tract digestibility and apparent ileal digestibility, as well as standardised ileal digestibility of essential amino acids and non- essential amino acids in weanling pigs [54]. The observed inconsistencies in results may be due to the variations in the amount, bacteria, KOH solubility, and enzymes used in each experiment.
GUT MICROBIOTA
An essential component of the gastrointestinal tract, the gut microbiome influences nutrition, physiology, and gut morphology. Furthermore, the microbiota plays a significant role in the host’s defense systems against infections [23,27]. FSBM has been shown to improve gut microbiota balance and address dysbiosis, by reducing the growth of pathogenic species while increasing beneficial bacteria, this may affect host health, performance, and disease risk [18]. The most widely utilized microorganisms in SBM fermentation include Lactobacillus, Streptococcus, Bacillus, Bifidobacterium, Enterococcus, Aspergillus, Candida, and Saccharomyces [27]. The gut microbiome is essential for immune, nutritional and protective functions, as it inhibits colonization by pathogens and the formation of harmful fermentation products [58]. As a result, the microbial diversity of the gut microbiome is associated with overall gut health. According to studies, dietary supplementation of FSBM could decrease coliforms and Salmonella, and improving intestinal microbial ecosystems [59]. Weaned pigs fed diets containing FSBM with starter cultures of B. subtilis, Hansenula anomala, and L. casei have been shown to increase Lactobacillus populations while decreasing E. coli [15]. Total anaerobic bacteria, lactic acid bacteria, Enterobacteria bacteria were shown to increase in distal small intestine, stomach, cecum, midcolon, as well as lactic, acetic, propionic, and butyric acid in the stomach of weaning pigs fed FSBM. This dietary inclusion promotes higher digestive enzyme activity, which improves feed utilization efficiency [46]. Additionally, an increase in the population of Lactobacilli and Bifidobacteria and a reduction of E. coli in jejunum, caecum and colon of piglets fed FSBM was reported [60]. Moreover, Qiu et al. reported a higher Lactobacillus and Faecalibacterium counts in the colon of weaned piglets fed FSBM diets. However, beneficial effects to tight junction proteins and inflammation in the colon were observed [19]. The supplementation of piglet diets with FSBM has been shown to impact the populations of the genus Lactobacillus, butyrate-producing bacteria (Clostridium, butyricum, Bacteroides, Butyrivibrio, and Eubacterium) and amino acid-fermenting bacteria (Enterobacteriaceae, Peptostreptococcus, Campylobacter, and Helicobacter) in the large intestine, thereby improving gut health [61]. Canibe et al. showed that dietary inclusion of FSBM in the diet of weaning piglets increased the population of lactic acid bacteria and Enterobacteriaceae, as well as an increase in lactic, acetic, propionic, and butyric acid in caudal small intestine, stomach, cecum, and midcolon compared to unfermented SBM [49]. Furthermore, a study conducted by Zhang et al. reported that the dietary inclusion of SBM fermented with L. plantarum and B. subtilis could increase the α-diversity of the gut microbiota, leading to an increase in the abundance of phylum Firmicutes, Bacteroidetes, and Bacteroidota, along with genera such as, Lactobacillus, Muribaculaceae, Ruminococcaceae, Prevotellaceae, and Rikenellaceae [20]. This also resulted in the reduction in the abundance of the phylum Proteobacteria and the genera Methanobrevibacter and Clostridium sensu stricto 1 in weaning piglets. The results showed that Rikenellaceae RC9 was positively associated with immunoglobulin G (IgG) and negatively associated with interleukin 1 beta (IL-1β), whereas the Prevotellaceae NK3B31 group was negatively associated with tumor necrosis factor-alpha (TNF-α) and urea nitrogen, and significantly associated with IgG, alkaline phosphatase and total protein. Muniyappan et al. reported that the dietary inclusion of SBM fermented with Enterococcus faecium in weanling pigs lead to an increase in the abundance of the phylum Firmicutes and the genera Lactobacillus, prevotella, Lachnospiraceae, and Lachnoclostridium. this inclusion also resulted in a reduction in the abundance of the phylum Bacteroidetes, Proteobacteria, and the genera Escherichia-Shigella, Clostridium sensu stricto1, Bacteroides, and Parabacteroides [18]. These three main populations of Bacteroidetes, Proteobacteria, and Firmicutes are crucial for growth and balance in energy metabolism. Similarly, supplementing SBM fermented with L. plantarum, B subtilis, and S. cerevisiae to the diets of weaning pigs can alter the microbial population and gut morphology [43]. Their results showed an increase in the phylum Firmicutes and the genera Lactobacillus and Prevotella, coupled with a decrease in the phylum Bacteroidetes and Proteobacteria and genera Escherichia, Streptococcus, and Stenotrophomonas in the cecum and colon. Species such as, Faecalibacterium, Rosepuria, and Pseudobutyrivibrio have high concentrations of butyrate in weaning pigs, which is crucial for gut cell metabolism [62]. The dietary inclusion of FSBM in the diet of weaning piglets resulted in an increased abundance of total bacteria, Lactic acid bacteria, and Clostridium perfringens in the caecal digesta, leading to an improvement of nutrient digestibility and enhanced gut morphology [51]. They found that dietary FSBM increased the levels of phylum Actinobacteria and Proteobacteria in faeces, as well as Firmicutes and Tenericutes in the colon of weaning pigs. Additionally, the genera Blautia, Coprococcus 2, Anaerostipes, Dorea, Bifidobacterium, and Roseburia were higher in the faeces of weaning pigs supplemented with FSBM, resulting in a reduced abundance of Bacteroides and Lachnospira in faeces. the genera Ruminiclostridium 5 and Lactobacillus were higher while Terrisporobacter and Clostridium sensu stricto 1 were lower in the colon of the weaning pigs fed FSBM compared to SBM. Positive correlations were observed between diarrhea in piglets and the relative abundance of Clostridium sensu stricto 1, Lachnospira, and Bacteroidetes [18]. This has been shown to have a positive effect on piglets growth performance and gastrointestinal health, by protecting the gut against infections and promoting efficient nutrient and energy extraction by Lactobacillus, Clostridium sensu stricto 1, and Blautia [63]. Immunoglobulin M (IgM) levels were shown to be positively associated with Lactobacillus and Lachnospira, while higher lymphocyte numbers were associated with Clostridium sensu stricto 1 [64]. Improvement in white blood cell counts were associated with elevated Blautia level [65]. In particular, Lactobacillus species, which are widespread and helpful in various parts of the digestive system, produce acetic acid, a compound that aids in reducing the abundance of the genus Campylobacter, which causes gastroenteritis, diarrhea and dehydration [66]. Moreover, Wang et al. showed that FSBM inclusion in the piglets diets increased the relative abundance of phyla Bacteroidetes, Acidobacteria, and Chloroflexi, and genus Prevotellaceae NK3B31 group, and resulted in a reduction in the relative abundance of phylum Proteobacteria and the genera Escherichia-Shigella, Clostridium sensu stricto 1, Campylobacter, and Actinobacillus in the cecal digesta compared to SBM when challenged with Enterotoxigenic E. coli [32]. These results are significant as lactic acid bacteria are known to positively affect the gut by regulating the gut microbiota makeup, creating immunity and increasing gut health [67].
GUT HISTOMORPHOLOGY
Efficient intestinal function is important for the development and welfare of weaning pigs. It not only protects against dangerous food chemicals and infections, but also plays important roles in digestion and the absorption of food nutrients, it additionally protects the sterile indoor environment from hostile light contents [23]. Some indicators of improving nutrient absorption include, increased measures of intestinal morphology, such as increased villus height, shorter crypt depth, higher villus height-to-crypt depth ratio. Additionally, the amount of goblet cells in the intestinal villi and crypts is also an indicator of intestinal health, as these cells produce mucin and prevent the adhesion of dangerous bacteria to the intestinal epithelium [68]. According to research, the villus height-to-crypt depth ratio is an important measure to assess the absorptive capacity of the small intestine in piglets, and as the villus height-to-crypt depth ratio increases, so does the amount of absorbed nutrients [43]. Piglets fed FSBM diets showed increased villus height (Duodenum: 432.7 μm and 417.3 μm; Jejunum: 595.7 μm and 478.2 μm; Ileum: 418.8 μm and 397.6 μm) villus/crypt ratio in the ileum, duodenum, and jejunum compared to those fed SBM, these improvements are concerned with promoting nutrient digestibility and growth performance [12]. Furthermore, Yang et al. reported that the dietary inclusion of FSBM increased villus height-to-crypt depth ratio in small intestine, which is positively related to feed intake [56]. Additional studies by Ma et al. showed that FSBM supplementation increased villus height (463 μm and 450 μm) and villus height-to-crypt depth ratio in the duodenum of weaning pigs [52]. In this study, improved morphology suggested that FSBM may reduce intestinal stress by preserving or improving the shape of the small intestine to enhanced absorptive capacity [52,55]. Yun et al. demonstrated that the supplementation of FSBM could increase villus height (Duodenum: 343 μm and 319 μm; Jejunum: 330 μm and 310 μm; Ileum: 318 μm and 301 μm) and villus height to crypt depth and reduce crypt depth (Duodenum: 214 μm and 222 μm) within the duodenum and ileum of weaned pigs [51]. Wang et al. also reported an improved villus height (Duodenum: 512 μm and 420 μm; Jejunum: 603 μm and 521 μm; Ileum: 412 μm and 315 μm) and villus height-to-crypt depth ratio, as well as reduced crypt depth (Duodenum: 280 μm and 320 μm; Jejunum: 280 μm and 395 μm; Ileum: 198 μm and 240 μm) in the ileum, duodenum, and jejunum of weaning pigs fed on FSBM diet [60]. Better nutrient absorption and reduced rate of tissue turnover is indicated by longer villi and less crypt depth [69]. Qiu et al. further reported that the dietary supplementation of fermented SBM with B. subtilis, B. licheniformis, L. plantarum, and Lactobacillus reuteri results in an increased villus height (Duodenum: 583 μm and 420 μm; Jejunum: 528 μm and 457 μm) and villus height-to-crypt depth ratio, and improved intestinal barrier function, possibly due to the response to intestinal inflammation [19]. Consequently, the lower concentration of antinutrients in FSBM may contribute to improved morphology.
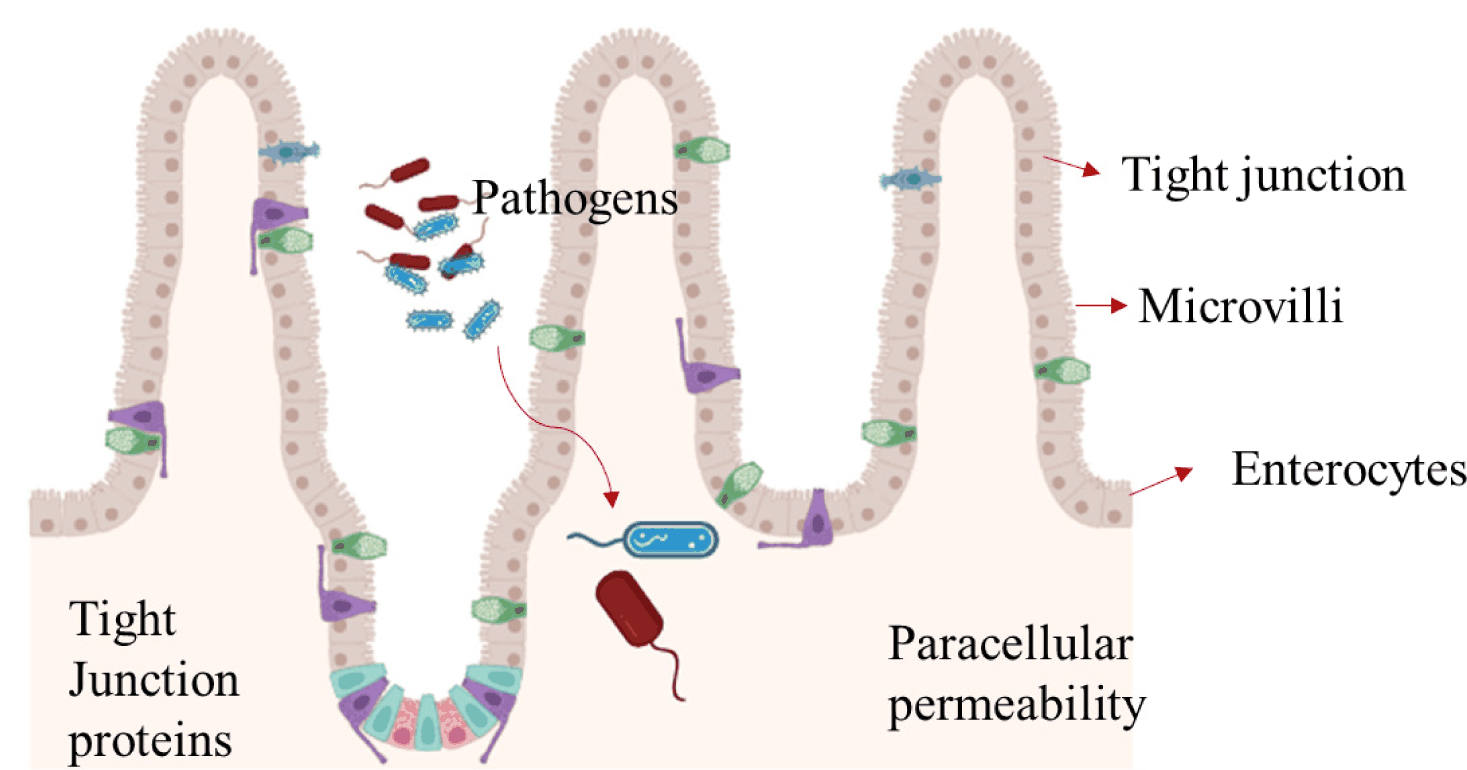
GUT BARRIER
The first line of defence in the digestive system is a complex mucus barrier, which is important for protecting the host from invading pathogens [70]. Intestinal epithelial cells, a monolayer in the gastrointestinal tract between the luminal and systemic portions of the gut, serve as the initial line of defence against pathogens and genotoxic chemicals. This epithelial barrier is formed by the combination of tight junction proteins and epithelial cells, preventing the passage of ions, toxins and microbes. A cell culture study by Zhang et al. demonstrated that SBM fermented with B. subtilis could significantly increase the survival rates and induce oxidative damage in the intestinal porcine epithelial (IPEC-J2) cells by regulating intracellular tight junction-related proteins in the jejunum and ileum of piglets [28]. The permeability of the intestinal barrier can become elevated as a result of loose junctions, making it easier for microbes and dangerous antigens to penetrate and cause intestinal inflammation [71]. Intestinal permeability can be measured by paracellular and transcellular flow of ions [72], and transepithelial electrical resistance (TER) serving as a reliable predictor of epithelial integrity. Thus, changes in TER and fluorescein isothiocyanate-dextran transit can indicate differential paracellular and transcellular permeability. The TER value is reduced in epithelial cells after FSBM significantly ameliorated E. coli K88-induced decreases [28]. Wang et al. showed that in FSBM, Enterotoxigenic E. coli K88 challenge can restore and maintain the intestinal mucosal barrier function of weaned pigs. Their findings also indicated that tight junction proteins such as, claudin-1, occludin, zonula occludens-1 (ZO-1), mucin 1 and mucin 2 can be more strongly expressed [32]. Additionally, Wang et al. reported a reduction in basal paracellular fluorescein isothiocyanate-dextran passage in the mid-jejunal segments of weaning piglets supplemented with FSBM challenged with Enterotoxigenic E. coli K88 [47].
TIGHT JUNCTION PROTEINS
A single layer of enterocytes and several protein complexes called “tight junctions” combine to form the intestinal barrier [72]. These tight junctions consist of many transmembrane and cytosolic proteins, such as occludin, claudins, zonula occludens, tricellulin, cingulin, mucin and junctional adhesion molecules, as well as intracellular plaque proteins such as, zonula occludens and cingulin [73]. The three primary structural components of tight junctions (TJs) are occludin, claudin, and ZO-1. These proteins are involved in the maintenance of intestinal integrity and barrier function [74]. Qiu et al. reported an increase in the mRNA expression levels of tight junction protein 1 (TJP1), claudin-1 and interleukin 10 (IL-10) and lower mRNA expression levels of interleukin 1β (IL-1β) and tumor necrosis factor α (TNF-α) in the duodenum, jejunum and colon of weaning piglets fed a FSBM diet supplemented with yeast, B. subtilis, B. licheniformis, L. plantarum, and L. reuteri [19]. Additionally, increased protein expression level of ZO-1 in duodenum, jejunum and ileum and increased protein expression level of claudin-1 in jejunum were observed. Key integral membrane proteins known as tight junction proteins have the ability to maintain barrier function by limiting the permeability of epithelial cells to low-molecular-mass molecules [75]. Moreover, research indicates that the gastrointestinal tract is more susceptible to E. coli invasion as a result of maternal stress, which causes the intestinal membrane to produce inflammatory factors and reduce the intestinal barrier integrity [76]. Zhang et al. reported that FSBM increased the mRNA expression of IL-1β, IL-8, and TNF-α by activating the phosphorylation of NF-κB, IκB-α, and p38 MAPK compared to the control Enterotoxigenic E. coli K88-treated IPEC-J2 cells [28]. Additionally, Zhang et al. reported that FSBM increased IL-10 and decreased interleukin 6 (IL-6) and interleukin 4 (IL-4) in the jejunum and ileum while resulting in lower tight junction proteins by activating the p38 MAPK signaling pathways [61]. Zhang et al. reported that FSBM supplementation increased IL-10 while IL-6) and interleukin 4 (IL-4) in jejunum and ileum and lowering tight junction proteins by activating the p38 MAPK signaling pathways [20]. These results are consistent with a previous investigation by Ma et al. which observed that FSBM supplementation significantly increased the mRNA expression levels of ZO-1 and occludin proteins in pigs, leading to an improvement in the intestinal barrier function [55]. The effects of fermented soybean meal intestinal barrier function and gut microbiota is shown in Fig. 2 and Table 3.
| Tight junction protein | SBM treatment (vs. SBM control) | Intestinal segment | Proposed mechanism | Observed change | References |
|---|---|---|---|---|---|
| Occludin | Bacillus-fermented FSBM (10% diet) | Jejunum | Microbial SCFAs enhance occludin transcription | ↑ 40%–50% protein expression | Zhang et al. [28] and Wang et al. [32] |
| Claudin-1 | Aspergillus-fermented FSBM (15% diet) | Ileum | Reduced TNF-α signaling | ↑ 30% mRNA stability | Li et al. [77] |
| ZO-1 | Mixed-culture FSBM (12% diet) | Duodenum | Probiotic-induced Protein kinase C activation | ↑ 25% phosphorylation | Zhang et al. [28] |
| Claudin-4 | Lactobacillus-fermented FSBM | Colon | Butyrate-mediated TJ assembly | ↓ 20% permeability | Qiu et al. [19] |
IMMUNITY
The organism’s internal environment is holistic, with changes in the microbiome composition influencing the immune responses of the animal [16]. FSBM has the potential to influence both mucosal and systemic immune responses of animals, thereby improving the overall health [31]. Primarily, FSBM suppresses inflammatory prostaglandins and produces nitric oxide, which may mitigate inflammatory activity, these anti-inflammatory properties have attracted considerable attention [23]. Among the cytokines that macrophages produces in response to FSBM exposure include, TNF-α, interleukin 12 (IL-12), interleukin 1β (IL-1β), interleukin 6 (IL-6), interleukin 2 (IL-2) and interleukin 10 (IL-10) [55]. In weaned pigs, it was reported that FSBM improved duodenal function and nutrient absorption, ultimately leading to enhanced immune status and growth [52]. The intestinal health and immunity of pigs can be improved by FSBM diet, which acts to inhibit the release of pro-inflammatory cytokines while stimulating immunoglobulins [56]. Zhang et al. reported decreased serum levels of IL-6, IL-1β and D-lactate of piglets fed FSBM supplemented diets [28]. Dietary FSBM may enhance pigs’ immune and intestinal health by promoting immunoglobulins and preventing the release of substances with pro-inflammatory properties [20]. Ma et al. showed that the dietary inclusion of FSBM could increase mRNA expression of Gpx1 and Gpx4 in liver while lowering the TNF-α concentration in the jejunum and duodenum of pigs [52]. In a recent study, FSBM supplementation was shown to regulate the inflammatory response and immune system in the jejunum and ileum, by inhibiting various signaling pathways such as, p38 MAPK, IκB-α, and p65 NF-κB in weaning pigs [61]. Furthermore, a study conducted by Yan et al. showed that the dietary inclusion of FSBM increased anti-inflammatory cytokine of IL-4 and decreased pro-inflammatory cytokines of IL-6 and IL-2 while increasing IgA, IgM, IgG and lowering diamine oxidase levels in the serum of weaning pigs [31]. Zhang and Piao. reported that the dietary inclusion of FSBM could decrease diamine oxidase concentration while endotoxin can promote the growth of beneficial intestinal barrier and gut microbes such as Lactobacillaceae and Lachnospiraceae [77]. Additionally, Wang et al. observed that Enterotoxigenic E. coli K88 challenged piglets had increased plasma D-lactate and diamine oxidase concentration while FSBM supplementation resulted in decreased plasma D-lactate and diamine oxidase concentration in the piglets [47]. The intestinal mucosal lesions and serum diamine oxidase correlate strongly; a greater value denotes more serious damage [78]. Under challenging conditions such as E. coli K88 prevalence, Wang et al. reported that fermented SBM supplementation with Streptococcus thermophiles, Saccharomyces cerevisae, B. subtilis and Aspergillus niger decreased plasma TNF-α, IL-1β, and IL-6 concentrations in weaning pigs [32]. Therefore, the improved intestinal barrier integrity may be related to the additional ability of FSBM to protect the intestines of weaned piglets from the inflammatory response. Meanwhile, intestinal inflammation is reduced by modifying the fermented soybean meal of intestinal microbes and their metabolites [79].
BLOOD BIOCHEMICAL PARAMETERS AND OXIDANT STATUS
The effect of FSBM supplementation on serum biochemical markers supporting the presentation of nutrient metabolism and physiological status in the body has been investigated by several authors [27]. Muniyappan et al. reported an increase in glucose, white blood cell, red blood cell and lymphocyte concentration as well as a decrease in the blood urea nitrogen concentration in piglets fed a diet of fermented SBM with Enterococcus faecium supplementation [18]. A reduction in the production of blood urea nitrogen concentration are better ways of protein catabolism, with its concentration and digestibility of amino acids and proteins been positively associated [18]. Furthermore, albumin, globulin, glucose, lymphocytes, lymphocytes ratio, white blood cell and phosphate have been shown to increase in weaning pigs as a result of the dietary inclusion of SBM fermented with B. subtilis and L. plantarum [12]. Xie et al. showed that the inclusion of SBM fermented with B. subtilis could improve glucagon concentration and lower the high-density lipoprotein cholesterol and creatinine concentration in pigs [13]. In another study, the dietary inclusion of B. subtilis and L. plantarum fermented SBM decreased triglyceride, total cholesterol, and blood urea nitrogen of piglets [20].
Fermented soybean meal supplementation has also been shown to increase the antioxidant status of pigs [80]. Zhang and Piao. reported an increased ferric reducing ability of plasma, total superoxide dismutase and glutathione peroxidase, alongside reducing malondialdehyde levels in weaning pigs fed diets supplemented with FSBM [77]. The dietary supplementation of FSBM could increase concentration of total superoxide dismutase, superoxide dismutase and glutathione peroxidase and reduce malondialdehyde levels in weaning pigs [52,55].
ENVIRONMENT IMPACT
A critical environmental problem in the pig industry is the production of harmful gases. Pig manure is a frequent source of greenhouse gas emissions that can result in great impact on to the environment. Methane, carbon dioxide, and nitrous oxide are just a few of the gases released by pig manure, with these gases been linked to global warming [81]. Several studies have demonstrated the deleterious effects of NH3 emissions on swine health and productivity. Furthermore, the environment is also affected by ammonia emissions from swine farms, as they cause eutrophication of surface water supplies and unpleasant odors [82]. FSBM has been shown to reduce harmful gas emissions in pigs through a complex multi-step, process. Firstly, Lactobacillus, prevotella, Lachnospiraceae and Lachnoclostridium are among the beneficial bacteria that can proliferate the gut of pigs fed a fermented SBM diet supplemented with Enterococcus faecium [18]. Cheng et al. reported a significant decrease in the NH3 levels, faecal nitrite and ammonia nitrogen of pigs fed SBM diets fermented with L. plantarum, B. subtilis, and S. cerevisiae [83].
Fermented soybean meal application alongside its major physiological responses in swine are presented in Table 4 and Fig. 3 respectively.
| Animal physiological status | Microbes | Response | Reference |
|---|---|---|---|
| Weaned pigs |
Lactobacillus plantarum, Bacillus subtilis, and Saccharomyces cerevisiae |
ADG ↑, G:F↓, diarrhea ↓, serum alkaline phosphatase, glucose, total protein, globulin, IgA, IgG, IgM, WBC, and lymphocytes ↑, blood urea nitrogen ↓. Duodenum, jejunum and ileum in Villus height and villus height to crypt depth ↑ and crypt depth ↓. |
Zhu et al. [12] |
| Weaning piglets | Bacillus subtilis BS12 | IL-6, IL-1β and D-lactate in serum ↓, ileum macrophage infiltration ↓. mucins 1, mucins 2, ZO-1, occludin, and claudin-1 in jejunum and ileum ↑. | Zhang et al. [28] |
| Weaned pigs | Enterococcus faecium SLB120 | DM, nitrogen, GE of AID ↑. | Jeong et al. [11] |
| Weaning pigs | B. subtilis CICC10088, B. subtilis CICC20030, B. subtilis CICC20076, B. subtilis CICC21076, and B. subtilis CICC23741 | ADG and G:F ↑, IL-4 and IL-6 ↓ and IL-10 ↑ in jejunum and ileum. | Zhang et al. [58] |
| Weaning piglets | Bacillus subtilis KC 101, Bacillus lactis RG 103, and Saccharomyces cerevisiae JM 102 |
Final BW, ADG, G:F ↑, digestibility of CP and total organic carbon ↑. carbohydrate-related metabolites of 4-aminobutanoate, 5-aminopentanoate, lactic acid, mannitol, threitol and β-alanine ↑. Protein catabolism of 1,3-diaminopropane, creatine, glycine and inosine ↓. |
Zhang et al. [33] |
| Weaned piglets | Bacillus subtilis, Bacillus licheniformis, Lactobacillus plantarum, and Lactobacillus reuteri | ADG, ADFI ↑, G:F ↓, diarrhea ↓, digestibility of CP crude fat ↑, ↑ duodenum and jejunum in Villus height and villus height to crypt depth, ↑ mRNA expression levels of duodenum and jejunum TJP1, claudin-1, IL-10 and ↓ mRNA expression levels of duodenum and jejunum TNF and IL1B, ↑ protein expression levels of duodenum and jejunum ZO-1 and claudin-1. Colonic levels of propionate and butyrate ↑, colonic levels of isobutyrate and isovelarate ↓. Genus of Faecalibacterium and Lactobacillus ↑. | Qiu et al. [19] |
| Piglets | NS | ADG and ADFI↑. Lactic acid bacteria and Enterobacteriaceae in cecum and midcolon ↑. Acetic, propionic, butyric acid in caudal small intestine, cecum and midcolon ↑. | Canibe et al. [44] |
| Weaned piglets | Lactobacillus plantarum NF8 | Final BW, ADG, ADFI ↑, G:F ↓. Duodenum, jejunum and ileum in Villus height and villus height to crypt depth ↑ and crypt depth ↓. ↑ Lactobacillus counts↓ Escherichia coli counts in jejunum, caecum and colon. | Wang et al. [57] |
| Weaned piglets | Lactobacillus plantarum IMAU80002, Bacillus subtilis CVCC717, and Saccharomyces cerevisiae w303 | ADFI, ADG ↑, Diarrhoea rate ↓. Lymphocytes, IgM, WBC in serum ↑. ↑ Phylum levels of Actinobacteria and Proteobacteria in faeces, and Firmicutes and Tenericutes in the colon. ↑ Genus level of Lactobacillus in faeces and colon. ↑ Family levels of Lactobacillaceae and Peptostreptococcaceae in faeces and colon. ↓ Family levels of Clostridiaceae, Lachnospiraceae and Bacteroidales S24-7 group in faeces and colon. ↑ Genera levels of Blautia, Anaerostipes, Coprococcus 2, Dorea, Roseburia, and Bifidobacterium in faeces. ↓ Genera levels of Lachnospira and Bacteoides in faeces. ↑ Genera levels of Lactobacillus and Ruminiclostridium 5 in colon. ↓ Genera levels of Clostridium sensustricto 1 and Terrisporobacter in colon. | Zhu et al. [84] |
| Weaned piglets | Enterococcus faecium SLB130 |
↑ BW, ADG, ADFI and G:F. ↓ Diarrhoea rate. ↑ DM, CP and GE. ↑ Glucose levels, WBC, RBC, and lymphocytes in serum. ↓ Blood urea nitrogen in serum. ↑ Alpha diversity. Phylum level of Firmicutes ↑ and ↑ Bacteroidetes, Proteobacteria. Genera level of Prevotella, Lactobacillus, and Lachnospiraceae ↑ and Lachnoclostridium and ↓ Escherichia-Shigella, Clostridium sensu stricto 1, Bacteroides and Parabacteroides. |
Muniyappan et al. [18] |
| Weaned pigs | Enterococcus faecium SLB130 | ↑ AID and ATTD of DM, CP, GE and amino acids. ↓ Urine nitrogen. | Muniyappan et al. [8] |
| Finishing pigs | Aspergillus oryzae GB-107 | ↑ ADG. ↑ Triglyceride in serum. ↓ Creatinine content in serum. Phylum level of Bacteroidetes ↑. Family levels of Prevotellaceae ↑. Order level of Bacteroidales ↑. Class level of Bacteroidia ↑. | Feng et al. [32] |
| Bamei Piglets | Lactobacillus plantarum QP28-1 and Bacillus subtilis QB8 | ↑ ADG and ADFI. ↑ IgM, IgG, total protein and IFN-γ in serum. ↓ TNF-α, IL-1β, IL-6, triglyceride, total cholesterol and blood urea nitrogen in serum. ↑ Alpha diversity. Phylum levels to Firmicutes and Bacteroidetes ↑ and Proteobacteria ↓. Genus levels of Lactobacillus, Muribaculaceae, Methanobrevibacter and Prevotella ↑ and Clostridium sensu stricto 1 ↓. | Zhang et al. [20] |
| Weaned pigs | Aspergillus oryzae GB-107 | ↑ G:F. ↑ histidine, lysine, and methionine digestibility. ↑ Blood urea nitrogen and total protein concentrations. | Cho et al. [29] |
| Weaned Piglets | Aspergillusoryzae 3.042 | ↑ ADG. ↓G:F. ↓ IgG and Lymphocytes in serum. ↓concanavalin A and lipopolysaccharide in serum and splenocytes. | Li et al. [77] |
| Weaned Piglets | Bacillus subtilis WB117 |
↑ ADG. ↓G:F. ↑total protease and trypsin activities in the duodenum and jejunum. Duodenum in Villus height and villus height to crypt depth ↑ and crypt depth ↓. Jejunum and ileum in Villus height ↑. |
Feng et al. [10] |
| Piglets | Lactobacillus casei CGMCC1.62, B. subtilis CGMCC1.504, and Hansenula anomala CGMCC2.881 | ↑ ADG and G:F. ↓ Diarrhoea rate. Digestibility of CP, crude fat, calcium and phosphorus ↑. Enzyme activity of protease, amylase and lipase ↑. ↑ Lactobacillus counts ↓ Escherichia coli counts | Yuan et al. [15] |
| Weaned pigs | Bacillus subtilis, Lactobacillus, and Saccharomyces cerevisiae | ↑ ADG and ADFI. AID and SID of CP and AAs ↑. ileal digesta digestive enzymes of Lipase, invertase and maltase ↑. Serum diamine oxidase ↓, T-AOC, SOD and GSH-Px ↑, MDA ↓. Immune function of IL-6, IL-2 ↓ and IL-4, IgA, IgM and IgG ↑. | Yan et al. [31] |
| Piglets | NS | ↑ BW, ADG, ADFI and ↓ G:F. ↓ Diarrhoea rate. ↑ Nutrient digestibility coefficients (ATTD and AID). ↑ Lactobacillus counts ↓ Escherichia coli and Clostridium perfringens counts in ileum and faeces. | Czech et al. [85] |
| Finishing pigs | Bacillus subtilis | ↑ Final BW and ADG. ↑ Digestibility of DM, CP, GE, ether extract and ash. ↓ High-density lipoprotein cholesterol and creatinine in serum. Antioxidant indices of CAT, T-AOC and SOD ↑ and MDA in serum and muscle ↓. ↑ Expression levels of the MyHC I and MyHC IIa in longissimus thoracis. | Xie et al. [13] |
| Weaned piglets | Lactobacillus, Bacillus, and Saccharomyces cerevisiae | ↑ Final BW, ADG, ADFI, and ↓G:F. ↓ Diarrhoea rate. Jejunum in Villus height and Villus height to crypt depth ↑. Ileum in Crypt depth ↓. ↑ α-Galactosidase, β-Galactosidase, α-Glucosidase and β-Glucosidase in cecum and colon. ↑ Propionate and Butyrate in cecum and colon. Phylum levels to Firmicutes ↑ and Bacteroidetes and Proteobacteria ↓ in cecum and colon. Genus levels of Lactobacillus and Prevotella ↑ in cecum and colon. | Xie et al. [37] |
| Weaning piglets | Lactobacillus acidophilus (BCRC10695), Lactobacillus delbrueckii (BCRC10696), and Lactobacillus salivarius (BCRC12574) | ↓ Diarrhoea rate. ↑ Lactobacillus counts ↓ Escherichia coli counts. ↑ IgG and IgM in serum. | Cheng et al. [22] |
| Nursery piglets | Lactobacillus plantarum (CGMCC No. 1.1209), Bacillus subtilis MA 139 and Saccharomyces cerevisiae (CGMCC No. 2.0707) | ↑ ADG and ADFI. ↑ Total protein, ↓ blood urea nitrogen in serum. ↓ Fecal ammonia nitrogen, Fecal nitrite and Fecal pH. Ammonia ↓. | Cheng et al. [82] |
| Sows and piglets | Aspergillus oryzae and Lactobacillus reuteri | ↑ Piglets BW. Oxidative stress status in GSH-Px, SOD and 8-iso-prostaglandin F2α ↑ and MDA and hydroxyl ion ↓ in sows serum. sows serum levels of estrogen, prolactin, IGF-1, and EGF ↓. |
GSH-Px, glutathione peroxidase; SOD, superoxide dismutase; MDA, malondialdehyde; IGF-1, insulin-like growth factor; EGF, epidermal growth factor; ADG, average daily gain; ADFI, average daily feed intake; G;F, feed to gain ratio; BUN, blood urea nitrogen; DM, dry matter; CP, crude protein; GE, gross energy; IgG, immunoglobulin G; IgM, immunoglobulin M; IgA, immunoglobulin; ATTD, apparent total tract digestibility; AID, apparent ileal digestibility; AA, amino acid; ; IL-6, interleukin 6; IL-1β, interleukin 1β; NS, not stated.
CONCLUSION
This review attempts to provide the most recent scientific findings regarding the use of FSBM in pig nutrition, underlining its beneficial effects on performance, digestibility, gut microbiota and morphology, immune response, and antioxidant status. The positive effects on gut health and function are largely determined by FSBM with low pH levels and soaring lactic acid bacteria and lactic acid concentrations. Therefore, FSBM can be used as a tactical tool in reducing the colonization by gastrointestinal pathogens in pigs. However, it is important to note that diets differed from one study to another in terms of substrate, microbial and fermentable material content, nutrient composition, and water content. Additionally, research has been conducted during critical phase across several global locations with varying environmental conditions. Therefore, the use of readily available fermented feed components locally, especially in poor countries, can reduce feed costs and guarantee viable broiler production. Notwithstanding, the lack of information on the use of fermented feed in pig production, future research should adopt a systematic approach and consider the effect of the components of the fermented feed on the microbiome of the pigs under investigation.