
INTRODUCTION
The negative perception of the livestock industry is increasing due to animal welfare and environmental concerns due to the increase in greenhouse gases and human population. Various alternatives to meat as sources of proteins, such as artificial meat derived from vegetables and the consumption of insects, have been suggested to solve these problems. However, the production of plant-based artificial meat is highly limited in use due to differences in its taste and texture compared to meat. Recently, the production of meat using cultured meat technology has emerged as an alternative to this problem. Cultured meat refers to edible meat obtained through cell proliferation without going through the process of raising livestock and is called in vitro meat or lab-grown meat. This concept of cultured meat has long been suggested, and stem cell technology has been actively applied in this area since its development. Founded in 2013 by Professor Mark Post of Maastricht University in the Netherlands, Mosa Meat succeeded in producing beef by cultivating stem cells; thus, the commercial use of cultured meat began in earnest. The use of cultured meat technology not only reduces the energy consumption for meat production to about 45% of that of existing methods of livestock breeding for meat but also reduces greenhouse gas emissions by more than 78%–96%, use of land by more than 99%, and use of water by 82%–96% compared to the use of animal livestock [1]. Moreover, it has the advantages of preventing or managing animal welfare issues and animal-derived diseases. However, there remain many obstacles to overcome to produce cultured meat for widespread consumption. In particular, since most of the growth factors and culture media used to produce cultured meat are derived from animals, cultured meat does not completely replace the use of livestock, and little is known regarding the safety of cultured meat consumption. Besides, it is difficult to clearly resolve this argue that the cultured meat might save the unethically slaughtered livestock since the producing cultured meat need animal sacrifice, suffering and death [2]. Another argue associated with producing cultured meat is that it is unethical to inform unhealthy foods even if it believe that the cultured meat will ethically be produce in the future [3]. In addition, the production of cultured meat is markedly more expensive than the use of conventional livestock; thus, there is a clear limit to its commercial use. Therefore, various attempts are currently being made to develop technologies that produce safe cultured meat inexpensively. At present, various laboratories worldwide are conducting research into cultured meat production, but only a few basic experimental methods have been reported regarding the production of cultured meat. Therefore, this study presents a basic experimental method for the production of cultured meat for commercial use and research (Fig. 1). This study is expected to assist with the production of cheap and safe cultured meat.
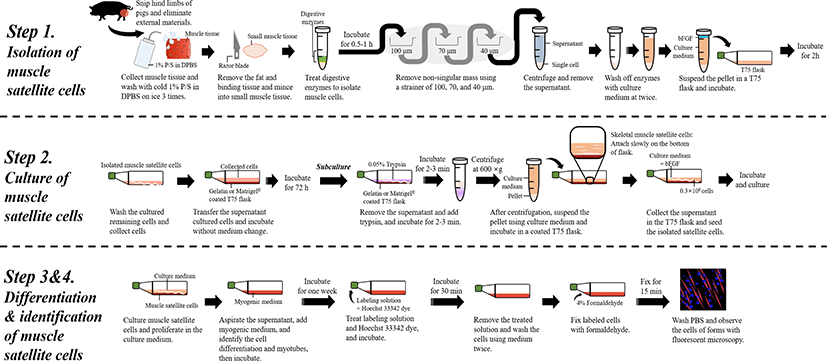
MATERIALS AND METHODS
Analytical grade isoflurane, ethanol (EtOH), bovine serum albumin (BSA), Triton X-100, paraformaldehyde, sucrose, sodium chloride, potassium chloride, sodium phosphate dibasic, potassium phosphate monobasic, 2-methylbutane, protease, and collagenase were purchased from Sigma-Aldrich (St Louis, MO, USA). Dulbeccos’ Modified Eagel’s Medium (DMEM), fetal bovine serum (FBS), horse serum (HS), and penicillin-streptomycin were purchased from Gibco (Thermo Fisher Scientific, Waltham, MA, USA). Anti-rabbit IgG (H + L) Alexa Fluor 488 conjugate was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA), and 4´,6-diamidino-2-phenylindole and dihydrochloride were purchased from Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA).
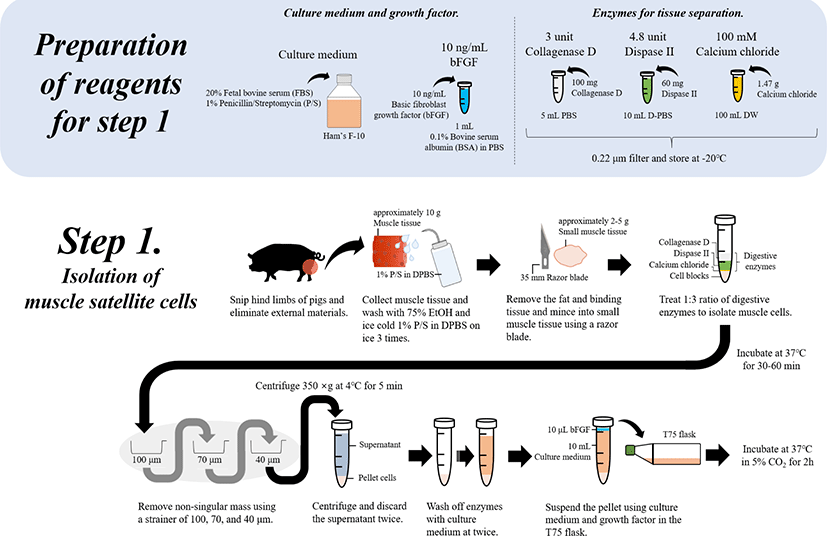
The cultivating medium for muscle cells was prepared with Ham’s F-10, FBS, and antibiotics (penicillin and streptomycin; P/S). Briefly, the final medium was prepared with 100 unit/mL of 20% FBS and 100 μg/mL of 1% 1X P/S and stored at 4°C. Myogenic medium for inducing differentiation was prepared with DMEM-high glucose (HG), HS, and antibiotics. Briefly, the final medium was prepared using 2% HS and 1% 1X P/S in DMEM-HG, and stored at 4°C until the end of use. The enzymes used for tissue separation were prepared by adding 3 units of collagenase D along with 4.8 units of dispase II. Finally, to achieve maximum collagenase activity, the digesting enzyme solutions were prepared with Dulbecco’s phosphate-buffered saline (DPBS) and filtered using a 0.22 μm filter (Whatman™, 9913-2502). These solutions were freshly prepared each time prior to digestion. Calcium chloride (1.47 g) was added to 100 mL of sterilized distilled water to form a 100 mM concentration, which was then filtered using a 0.22 μm filter and stored. The 10 ng/mL basic fibroblast growth factor (bFGF) solution was prepared with 10 ng of bFGF by adding 1 mL of 0.1% BSA to PBS. A total of 100 mL of 75% EtOH was prepared using 100% EtOH (75 mL) and sterilized ultra-pure water (25 mL). Sterile single-use disposable plastic wares, such as cell strainers (40, 70, and 100 μm), petri dishes, polypropylene centrifuge tubes (15 and 50 mL), and T-flasks (T75), were used. A clean environment was maintained that included a water bath, centrifuge, incubator, and microscope.
-
Euthanize the animal (pig) according to the institute’s animal care guidelines, and clean the surface of desired muscle region with EtOH spray. Then, resect the muscle (from the hind limbs) of the animal using surgical tools.
-
While collecting the muscle, take care to ensure that hair and any other external materials are not included.
-
Sterilize the collected (approximately 10 g) muscle tissue (approximately 1 × 1 cm) with 75% EtOH and wash three times with ice cold 1% P/S in DPBS (v/v).
-
Remove the fat and other binding tissues, and mince the muscle tissues (to approximately 2–5 g and 5 × 5 mm) using sterilized scissors or a razor blade (35 mm).
-
To isolate muscle cells, use a 1:3 ratio of the prepared digestive enzyme solution for the digestion of the external intercellular matrix by combining the cells and solution in a 15 mL tube at 37°C for 30–60 min.
* To prepare digestive enzymes, mix the digestive enzymes following 1 mL of 3 U collagenase D (final concentration: 1.5 U collagenase D), 1 mL of 4.8 U dispase II (final concentration: 2.4 U dispase II) and 50 μL of 100 mM calcium chloride (final concentration: 2.5 mM calcium chloride). Generally, several types of enzymes can be used to digest muscle tissues, such as proteases, collagenases (e.g., collagenase VI), trypsin, and pronase, which can affect the extracellular matrix.
-
After enzymatic digestion, remove non-singular mass using a strainer of pore sizes 100, 70, and 40 μm.
-
Recover the filtered cells by differential centrifugation (350×g for 5 min at 4°C). Discard the supernatant and repeat this step twice to wash off enzymes and condition the cells by further digestion and pH alterations using culture medium with 20% FBS.
-
Recover the pelleted cells (approximately 1 g) and suspend as 10 mL of culture medium (20% FBS in Ham’s F-10 with 1% P/S) and 10 ng/mL bFGF in a T75 flask.
* bFGF promotes satellite cell proliferation and inhibits the differentiation of myoblasts [4]. In addition, some studies have suggested that the addition of leukemia inhibitory factor, transforming growth factor-β, and insulin-like growth factor-I can improve animal muscle satellite [5,6].
-
Incubate the suspended cells in the T75 flask at 37°C in a 5% CO2 incubator for 2 h. Therefore, this step can be applied to isolate whole muscle cell populations with many satellite cells from animals (Fig. 2).
Further steps were carried out to culture the isolated muscle cells to produce cultured meat.
-
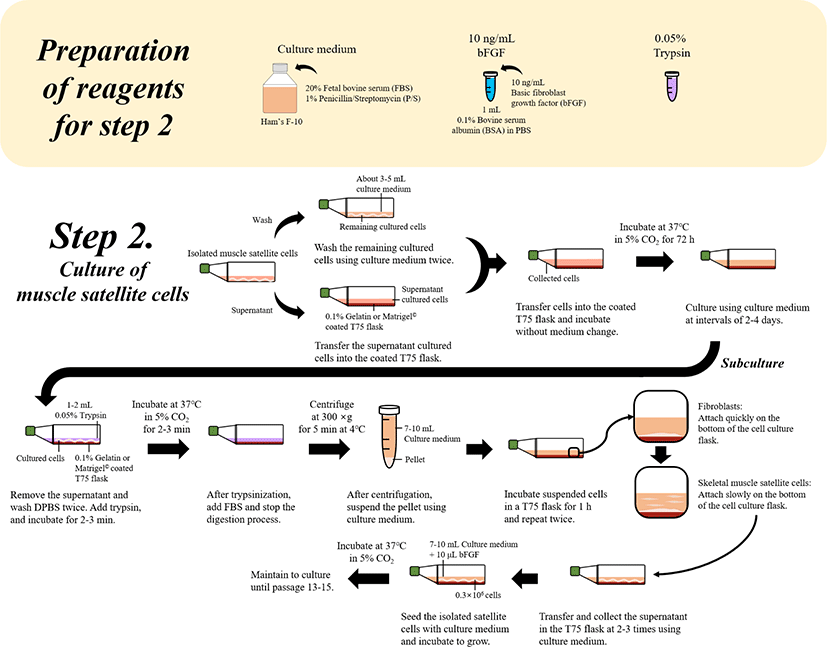
Transfer the supernatant obtained following the above steps (isolation of muscle cells) in the incubated T-75 flask into either a 0.1% gelatin- or Matrigel©-coated T75 flask.
-
Collect the remaining cultured cells (No. 9) in the T75 flask using culture medium (approximately 3–5 mL) twice and then transfer into a new T75 flask.
-
Incubate the transferred cells in the gelatin- or Matrigel-coated T75 flasks without medium change at 37°C in a 5% CO2 incubator for 72 h.
-
After 72 h, wash the incubated cells and change the culture medium at intervals of two to four days.
-
For subculturing, remove the old medium and wash with DPBS twice and add 1–2 mL of 0.05% trypsin, followed by incubation at 37°C in a 5% CO2 incubator for 2–3 min.
-
After incubation, to maintain cell viability, add the 0.5 mL of DPBS to neutralize trypsin and stop the digestion process. From the neutralized trypsin solution, separate the cells using a centrifuge at 300×g for 5 min at 4°C.
-
After centrifugation, suspend the pellet in 7–10 mL of culture medium (20% FBS in Ham’s F-10 with 1% P/S).
-
Incubate the suspended cells in 0.1% gelatin-coated T75 flasks for 1 h, repeating twice to separate only the fiber cells and sample muscle satellite cells.
* While fibroblasts and skeletal muscle satellite cells were cultured in 0.1% gelatin-coated T75 flasks due to different culture characteristics, fibroblasts quickly attached to the bottom of the cell culture flask and muscle satellite cells were slow to attach and remained in the suspended fluid [7].
-
Based on the characteristics of muscle satellite cells, collect the supernatant as satellite cells (approximately 7-10 mL) in the T75 flask and transfer into a 0.1% gelatin-coated T75 flask.
-
This step was repeated two to three times using culture medium (20% FBS in Ham’s F-10 with 1% P/S) to collect the remaining cells, which might have been rich in satellite cells.
-
Seed the isolated satellite cells at a density of 0.3 × 106 cells with 7–10 mL of culture medium (20% FBS in Ham’s F-10 with 1% P/S and 10 ng/mL bFGF).
-
Incubate the cells to grow at 37°C in a 5% CO2 incubator.
-
Maintain the cells by subculturing until that passage number of cells is 13–15 (Fig. 3).
-
To induce the differentiation of muscle satellite cells, change the culture medium to myogenic medium (2% HS and 1% P/S).
-
Aspirate carefully the cultivation medium, and add the cultured muscle satellite cells to 10 mL of myogenic medium (2% HS and 1% P/S) and then incubate at 37°C and 5% CO2 for one week.
-
After one week, observe the differentiation of cells and formation of myotubes using fluorescent dye reagents (Fig. 4).
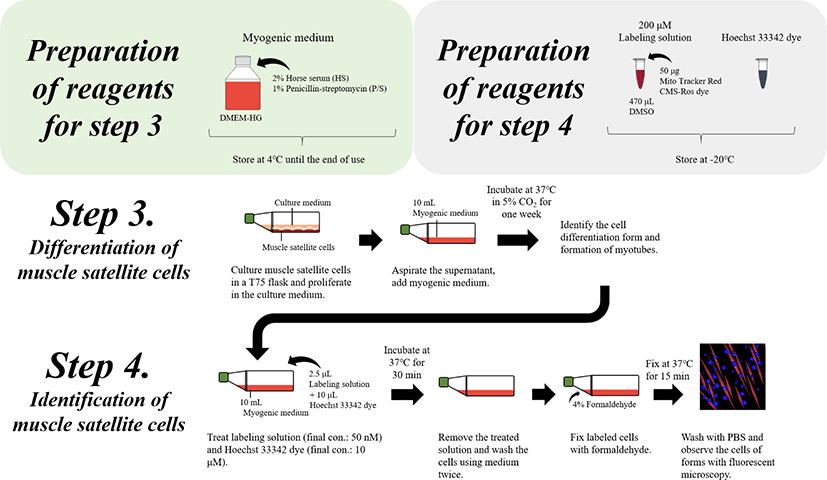
The biomarkers of satellite and differentiated cells were used to identify the satellite and differentiated cells as well as myotube formation as indicators of myogenesis. Here, the differentiation cells were quantified using the cell fusion index, which refers to the percentage of nuclei inside myotubes compared to the total count of nuclei in the sample [6]. Therefore, the fusion index was assessed by determining the visualization of nuclear staining, such as by using fluorescent dyes (e.g., bisBenzimide H 33342 trihydrochloride and Mito Tracker Red CMX-Ros) [8] and Giemsa staining.
-
Prepare the fluorescent dye solution (200 μM of Mito Tracker Red CMX-Ros) from the stock right before staining the cells and immediately use as a fresh preparation to avoid the loss of fluorescence ability.
* Prepare a 200 μM fluorescent dye stock solution by dissolving 50 μg of Mito Tracker Red CMX-Ros dye in 470 μL of DMSO, which is then stored at −20°C.
-
Treat the differentiated cells in the T75 flasks with 10 mL of medium with labeling solution (2.5 μL of 200 μM Mito Tracker Red CMX-Ros [final concentration: 50 nM] and 10 μL of Hoechst 33342 dye [final concentration: 1 μM]) for 30 min at 37°C.
-
After incubation, remove the labeling solution and Hoechst 33342 dye, and wash the cells using cell medium twice.
-
Fix the Labeled cells with 4% formaldehyde for 15 min at 37°C, followed by washes in DPBS.
-
Observe differentiated cells or myotubes using fluorescence microscopy (Fig. 4).