
INTRODUCTION
Efficient and sustainable production of meat from broiler chickens relies heavily on the ability of the animal to digest and absorb essential nutrients. Minerals such as zinc (Zn), copper (Cu), iron (Fe), manganese (Mn), and selenium (Se) are indispensable for growth, development, and overall health [1,2]. The ingredients and sources of these minerals, including inorganic, organic, and nano-forms, profoundly affect their digestibility and subsequent utilization by livestock species [3,4]. The choice of mineral sources in broiler chicken diets is a topic of ongoing debate and research in animal nutrition. Although each type of mineral source has advantages and disadvantages, organic and nano-trace minerals are often considered superior to inorganic minerals for several reasons, such as bioavailability, the nature of antagonistic interactions, and environmental impacts [5,6]. Organic minerals are bound to organic molecules such as amino acids, peptides, or polysaccharides, which can enhance their absorption in the digestive system [5], whereas nano-sized minerals have a larger surface area, potentially leading to increased absorption [7]. This improved bioavailability indicates that a higher percentage of the mineral is absorbed and utilized by animals, reducing the risk of mineral waste and the need for excess supplementation [7]. Research has also shown that inorganic minerals compete with each other for absorption in the digestive tract [8,9]. For example, excess Cu can interfere with the absorption of minerals, such as Zn and Fe.
Organic and nano-minerals, which are more easily absorbed, may mitigate these antagonistic interactions, ensuring that poultry can access a broader spectrum of essential minerals without hindrance [7,8]. They are generally excreted in lower amounts in feces (5.77%), reducing the environmental impact of excess mineral discharge into the soil and water systems [7]. This could contribute to sustainable and eco-friendly livestock production practices. However, nanoparticles have a larger surface area, potentially leading to enhanced absorption in the gastrointestinal tract [10,11]. Thus, mineral engineering at the nanoscale level represents a cutting-edge approach to improving mineral digestibility. However, the safety and long-term effects of nano-sized minerals on animal nutrition remain a subject of ongoing research and debate. It is also important to note that the availability of trace minerals in basic feed ingredients may vary widely depending on factors such as soil quality, geographical location, and farming practices. While there is a growing body of research exploring the effects of different mineral sources on the digestibility of broiler chickens, several knowledge gaps remain in understanding how different mineral sources affect digestibility for sustainable and efficient livestock production in a world with an ever-increasing demand for animal proteins. The objective of this study was to evaluate the availability of trace minerals in feed ingredients and supplemental sources for broiler chickens.
MATERIALS AND METHODS
This study was conducted at the animal metabolism facility of Kangwon National University in Chuncheon, Korea, and all protocols used in this study were approved by the University’s Institutional Animal Care and Use Committee (Approval No: KW-180907-1).
The trace mineral premix used in this study were prepared in three different types such as inorganic, organic, and nano. The inorganic premix consisted of sulfate monohydrate form in Cu (CuSO4 H2O; 34% Cu), Fe (FeSO4 H2O; 30% Fe), Zn (ZnSO4 H2O; 35% Zn), and Mn (MnSO4 H2O; 31% Mn), and sodium form in Se (Na2SeO3; 43% Se). Amino chelate minerals were used as the organic premix, composing Cu (20%), Fe (20%), Zn (20%), and Mn (20%), and Se yeast containing 10% of Se was used in the organic premix. Nano-trace minerals were prepared by hot-melt extruder processing (HME), which was explained by Lee et al. [7]. The nanoparticle sizes of Cu, Fe, Zn, Mn, and Se showed 84, 97, 99, 104, and 107 nm on average, respectively. All inorganic and organic minerals used in this study were purchased from TMC(Anyang, Korea), and Sel-Plex® (Alltech, Nicholasville, KY, USA) was used as the Se yeast.
Prior to the experimental period, all chicks were fed a commercial starter diet from d 1 to 7 and grower diet from d 7 to 15 days so that they had normal body conditions with similar weights among the experimental treatments. At d 15 of age, a total of 480 birds, one-day-old Ross 308 males, were used in the current trial and housed in metabolic cages for chickens (0.8 × 0.9 m). All birds were randomly arranged according to their body weight (436 ± 23 g) and allotted to 8 experimental diets in a completely randomized design. There were 12 replicates in each diet group with 5 birds per replicate. The experimental diets consisted of 7 semi-purified diets, containing corn, soybean meal (SBM), corn gluten meal (CGM), fish meal, inorganic premix, organic premix, and nano-premix. An M-free diet was used to determine the basal endogenous loss of trace minerals loss, and all test diets were formulated to meet or exceed the requirements of crude protein, amino acids, Ca, and P according to Aviagen [12] (Table 1). Titanium dioxide (TiO2) at 0.3% was supplemented as an indigestible marker to determine the ileal digestibility of trace minerals.
Supplied per kilogram of diet: 9,000 IU vitamin A; 1,800 IU vitamin D3; 30 mg vitamin E; 1 mg vitamin K3; 1 mg vitamin B1; 10 mg vitamin B2; 4 mg vitamin B6; 0.02 mg vitamin B12; 12 mg pantothenic acid; 30 mg niacin; 0.20 mg biotin; 0.50 mg folic acid.
All birds were raised in metabolic cages equipped with a couple of nipples and a hopper feeder per cage, facilitating access to water and feed ad libitum. Birds were kept at temperature of 34°C for 3 days, and thereafter, the room temperature was gradually decreased by 3°C per week until it reached 24°C.
Birds were fed their assigned experimental diets from days 15 to 26 of age, which contained an adaptation period for 7 days and sampling periods for 5 days [10]. From 22 to 26 days, trays covered with plastic were placed underneath pens to collect excreta samples. Excreta samples were collected two times a day, and the samples from each of pens were pooled in a tray and stored at −20°C until required for analysis.
The excreta and feed samples were dried in a forced-air oven at 60°C for 72 hours and grounded in a Wiley Mill (Thomas Model 4 Wiley Mill, Thomas Scientific, Swedesboro, NJ, USA) using a 1-mm screen. The grounded samples (about 1 g) were weighted and heat-treated at 600°C for 1 hour in an electric muffle oven. The ashed samples then were cooled and lysed with 10 ml of 50% HCl (v/v), and were kept covered overnight. The samples were filtered using Whatman filter paper into a 100 mL flask known as a volumetric flask, which was washed two to three times, after which the samples were diluted with deionized distilled water. For the plasma samples, 1 mL samples were measured in porcelain crucibles and oven-dried for 4 hours at 105°C and then ashed for 1 h at 600°C in a muffle furnace. The trace mineral contents of the feed, ileum, and excreta were determined by inductively coupled plasma emission spectroscopy according to the methods of AOAC [13].
AID and SID of mineral was calculated according to the below equation described by Jeon et al. [14].
Mdiet and Mdigesta are mineral content in the diet and ileal output, respectively (mg/kg of DM [dry matter]); Crdiet and Crdigesta are chromium content in the diet and ileal digesta, respectively (mg/kg of DM); and IMend refers to the basal ileal endogenous loss of an mineral (mg/kg of DM intake).
The effects of dietary mineral supplementation (inorganic, organic, and HME) and mineral source (corn, SBM, CGM, fish meal) were determined using a one-way ANOVA procedure (SAS Institute, Cary, NC, USA). The difference of means was tested by the Tukey test. A significant difference was expressed in either p < 0.01 or p < 0.05, however, p values 0.05 to 0.1 were given to indicate if the value tended to differ.
RESULTS
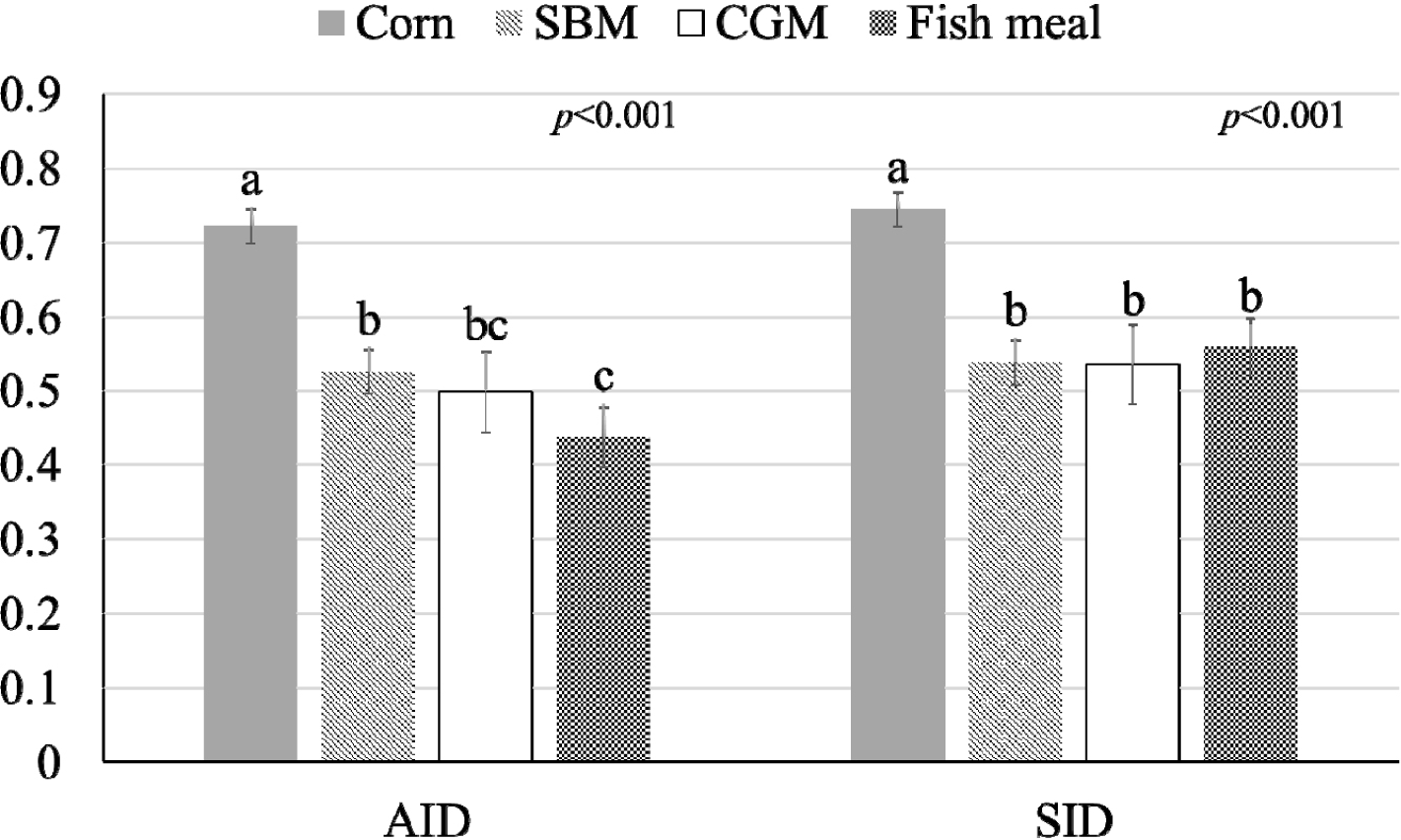
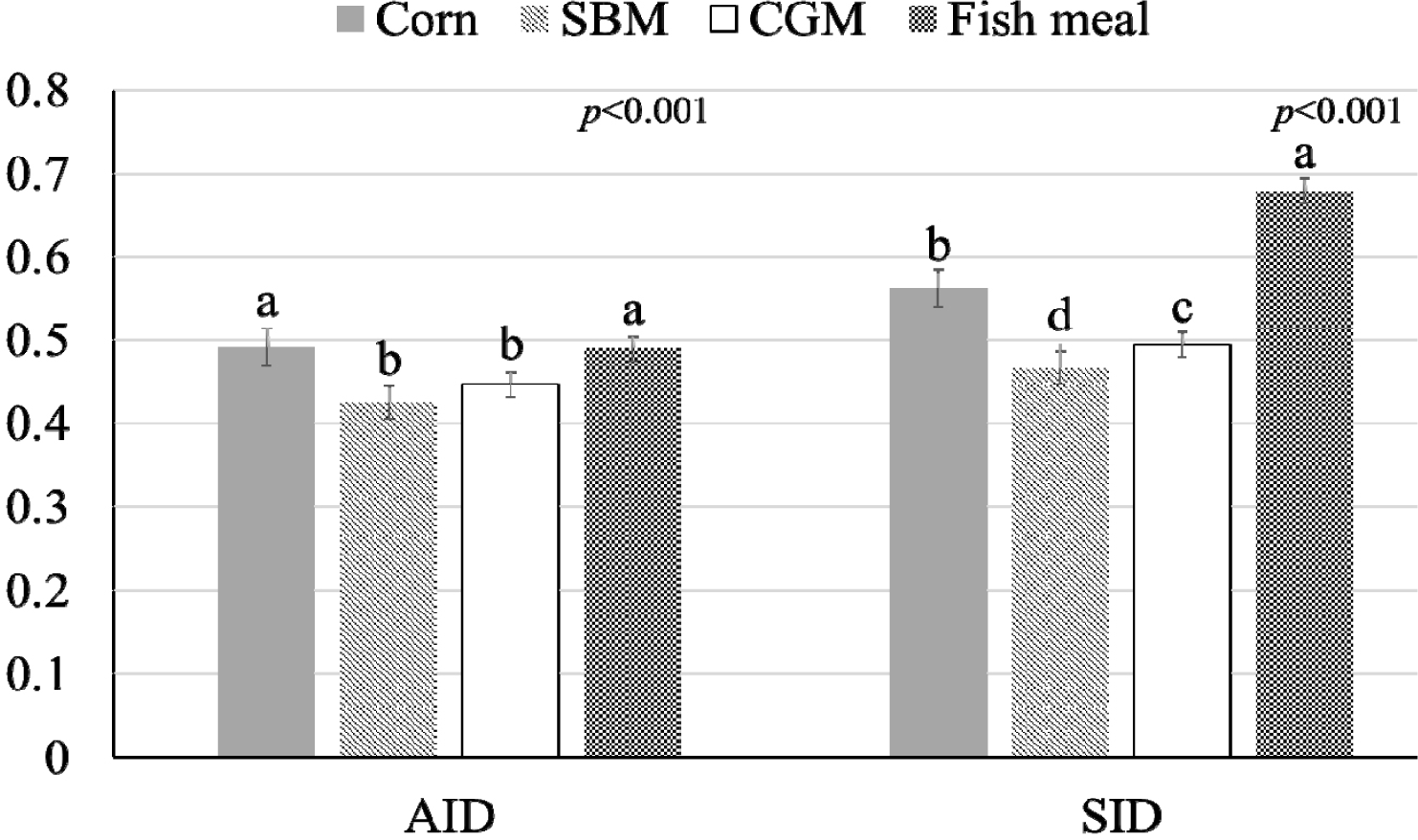
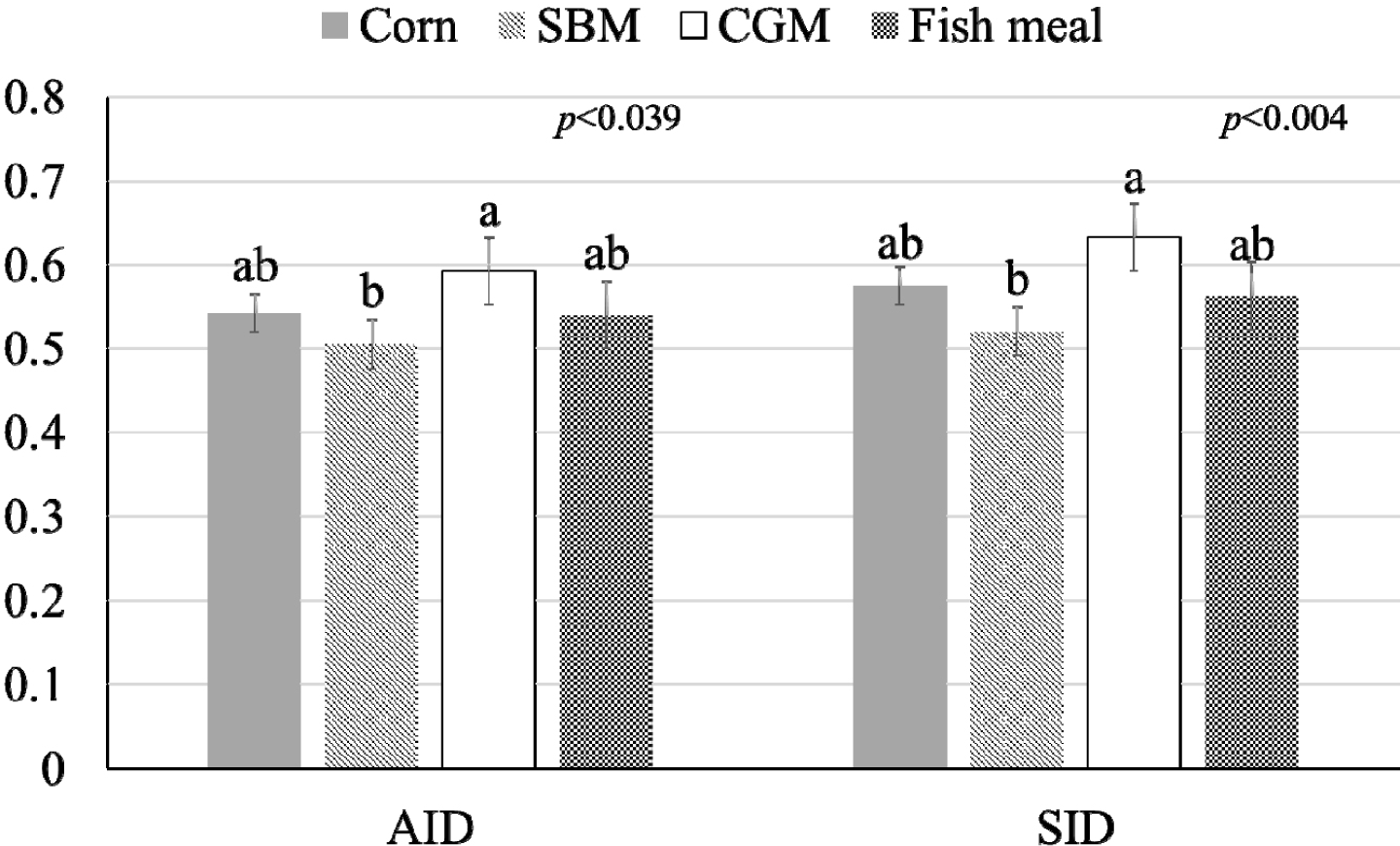
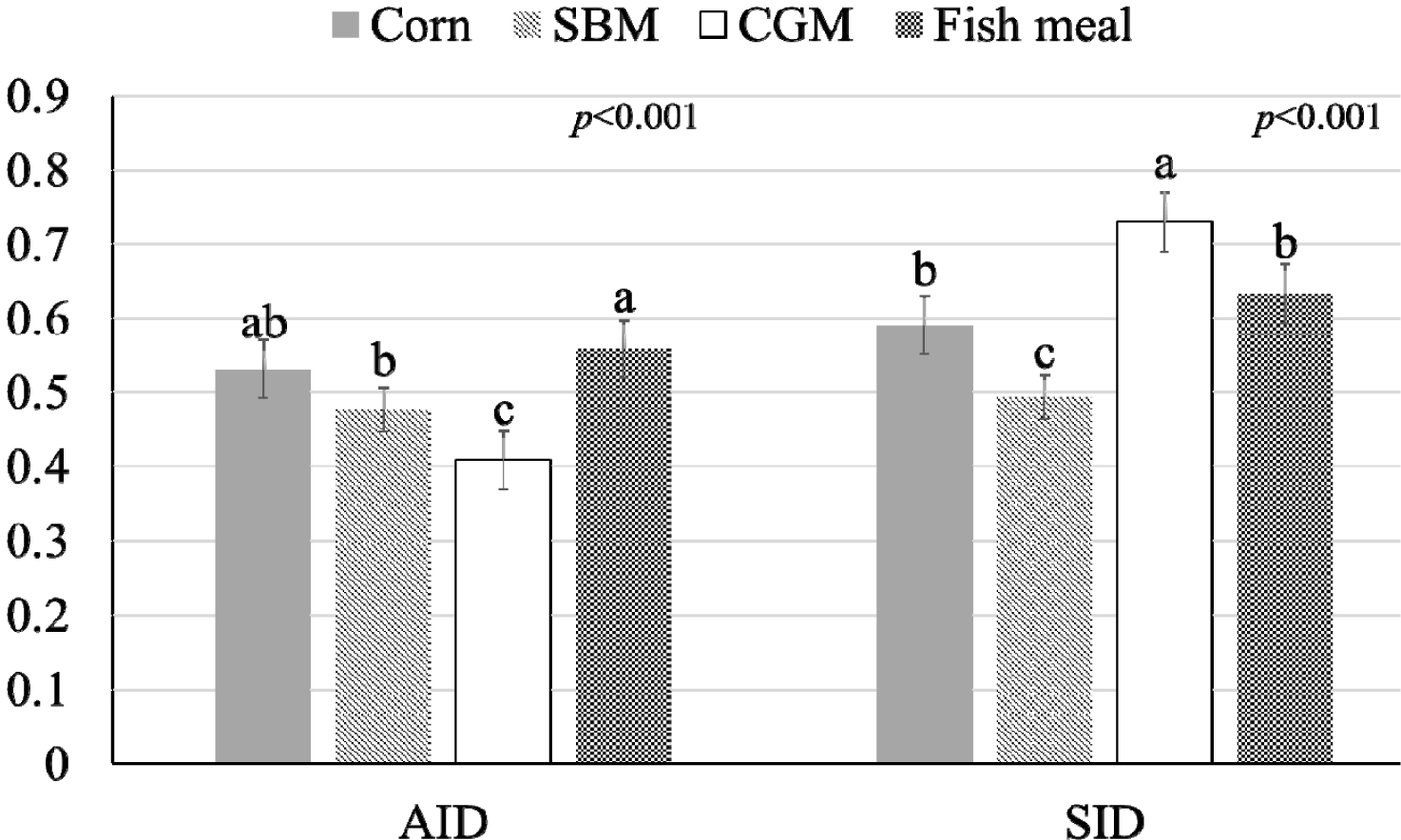
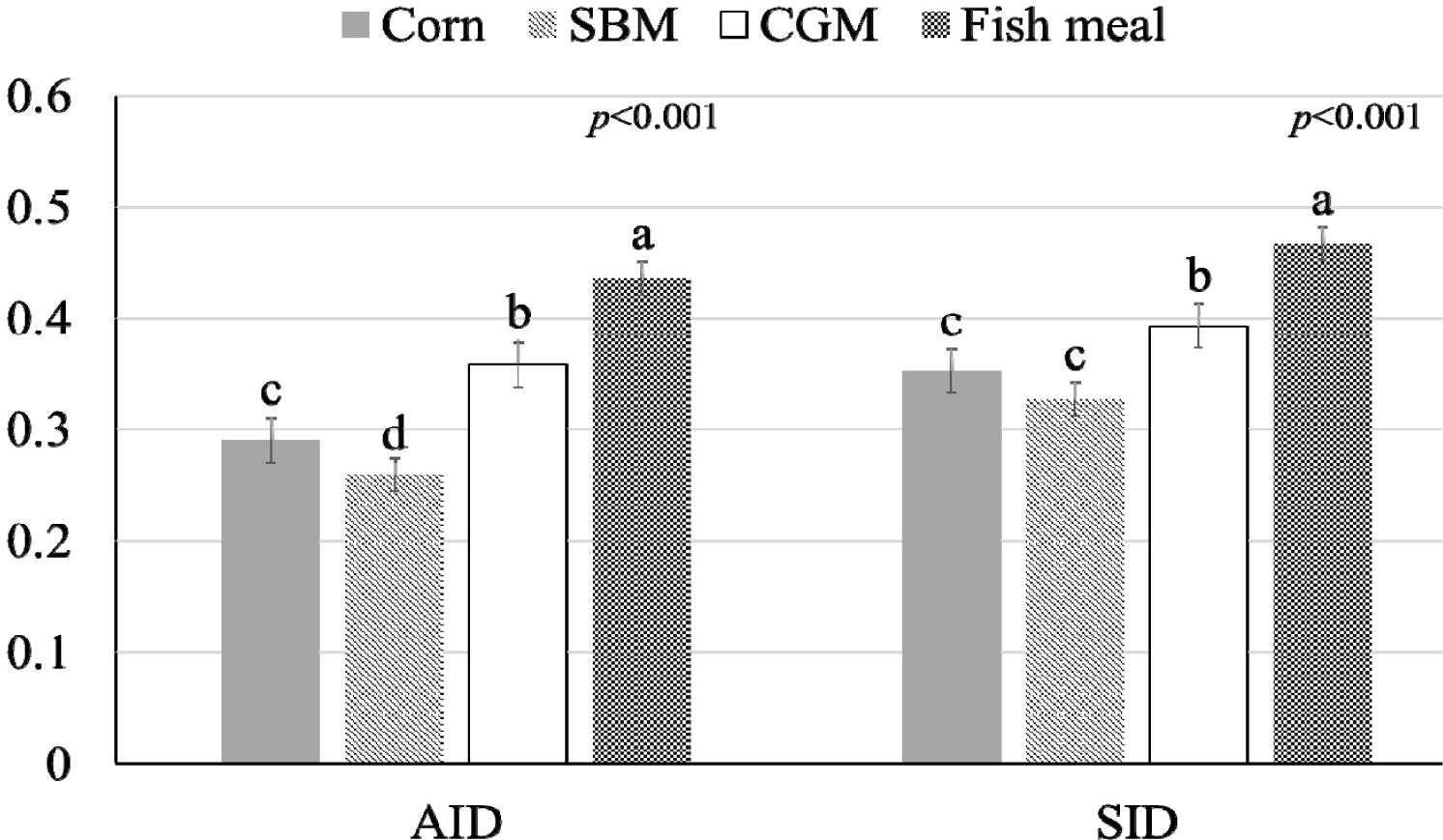
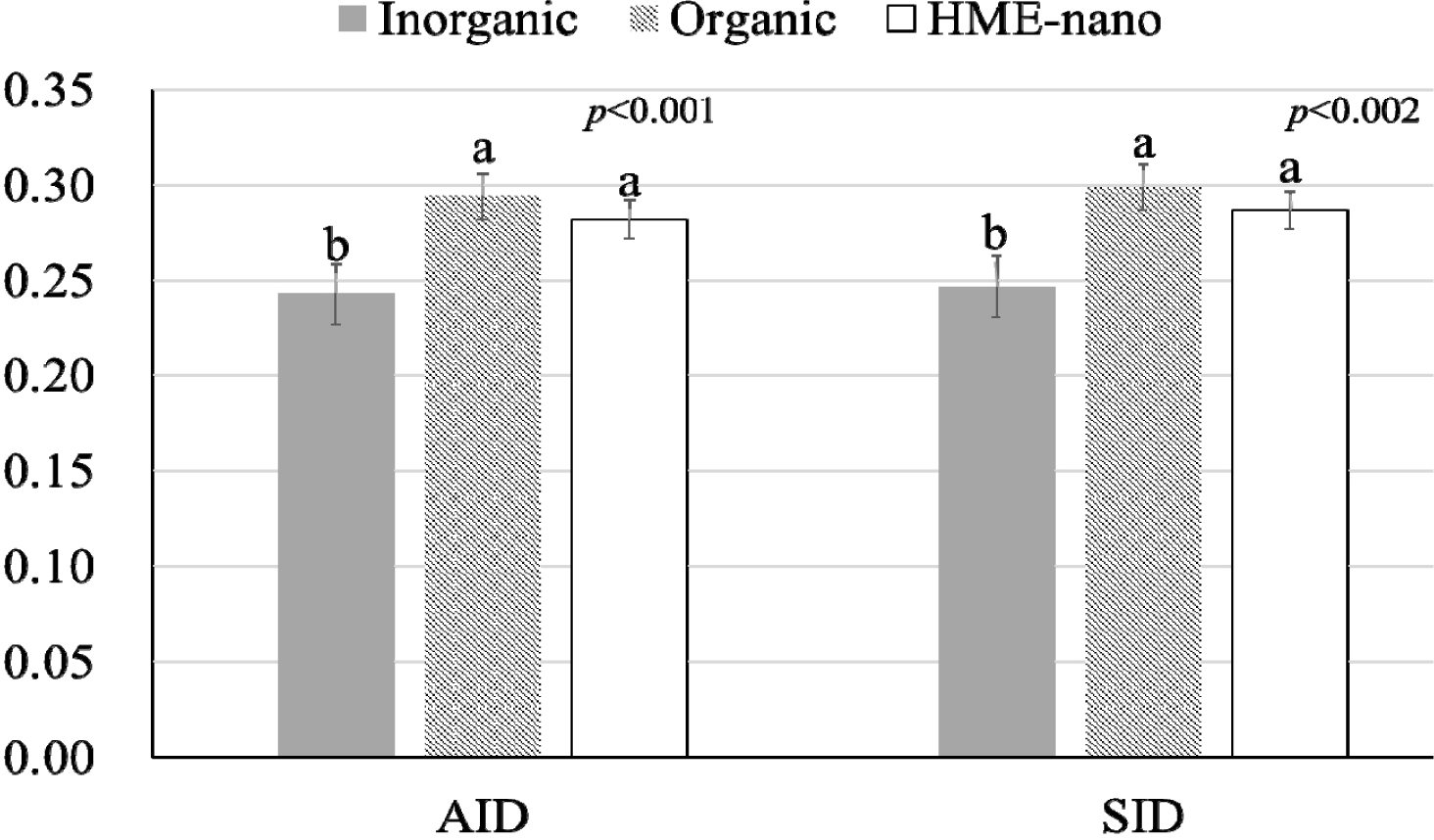
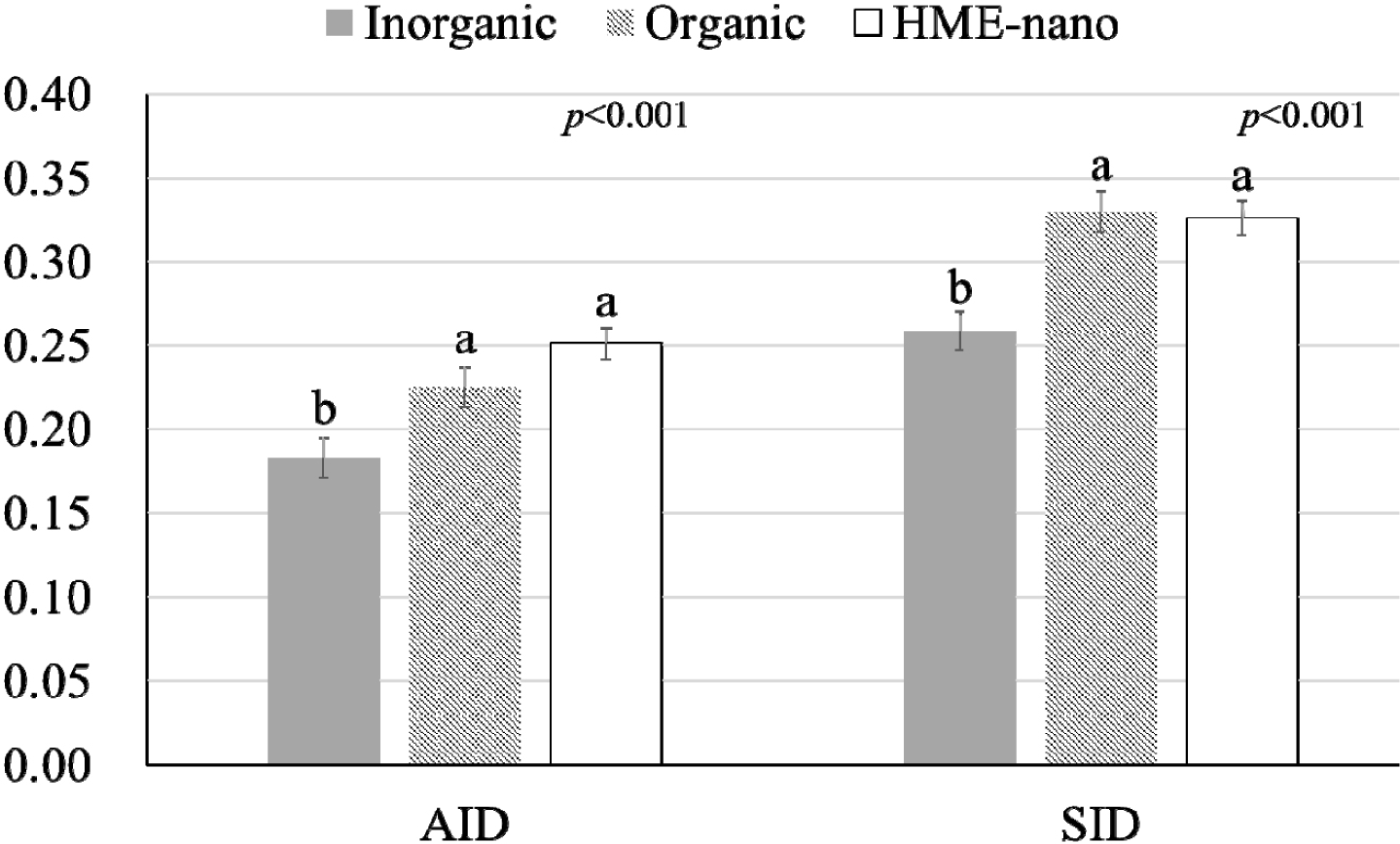
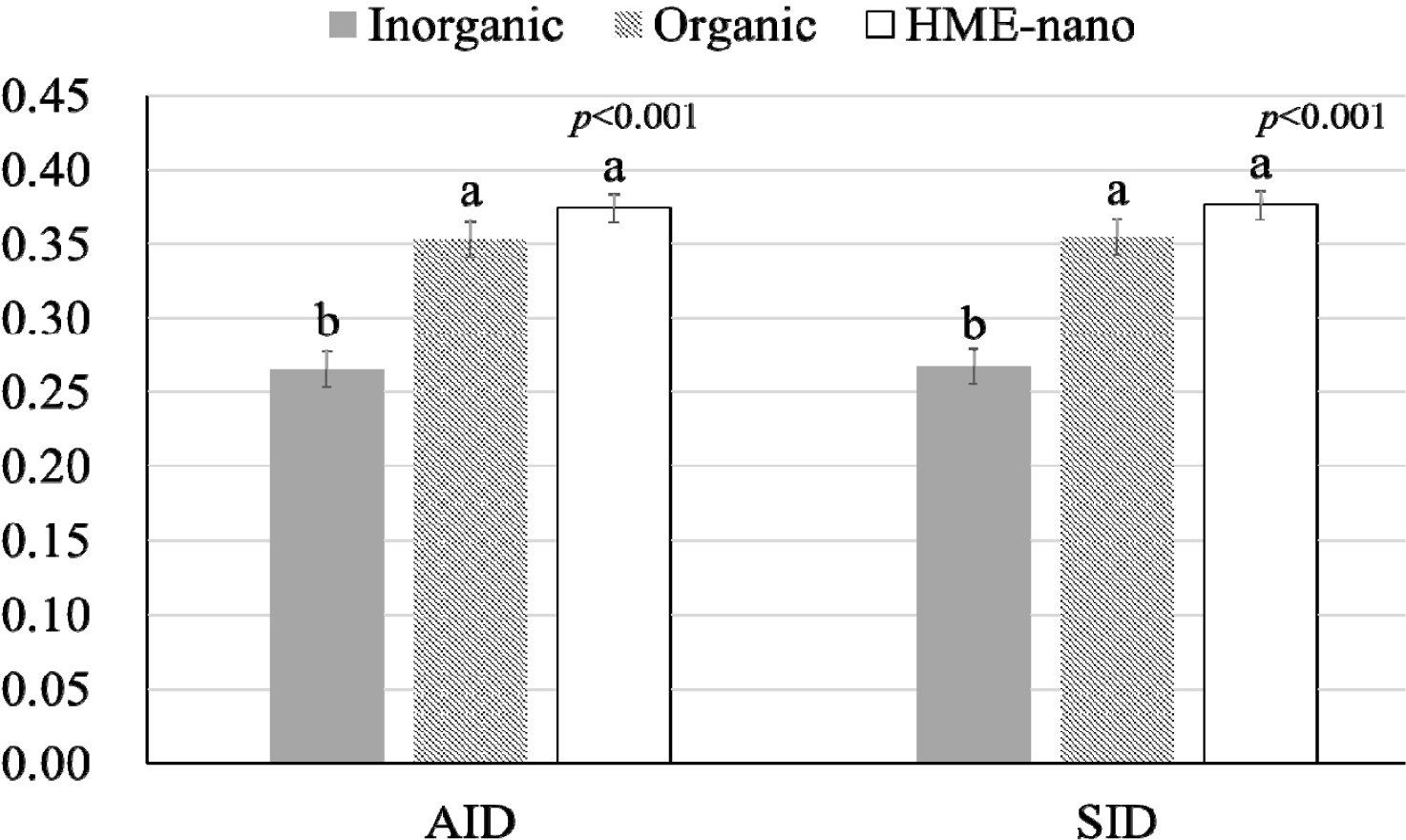
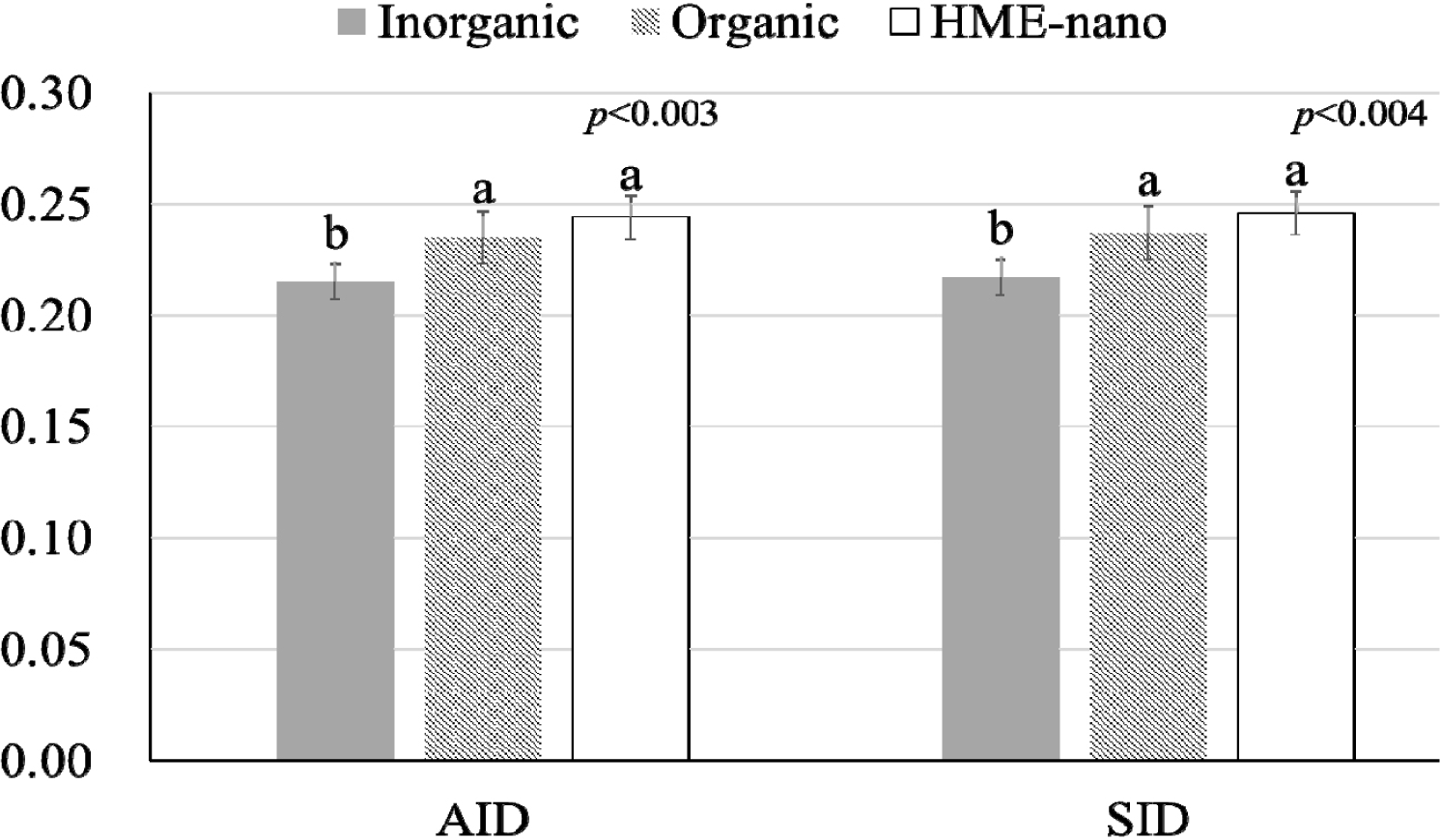
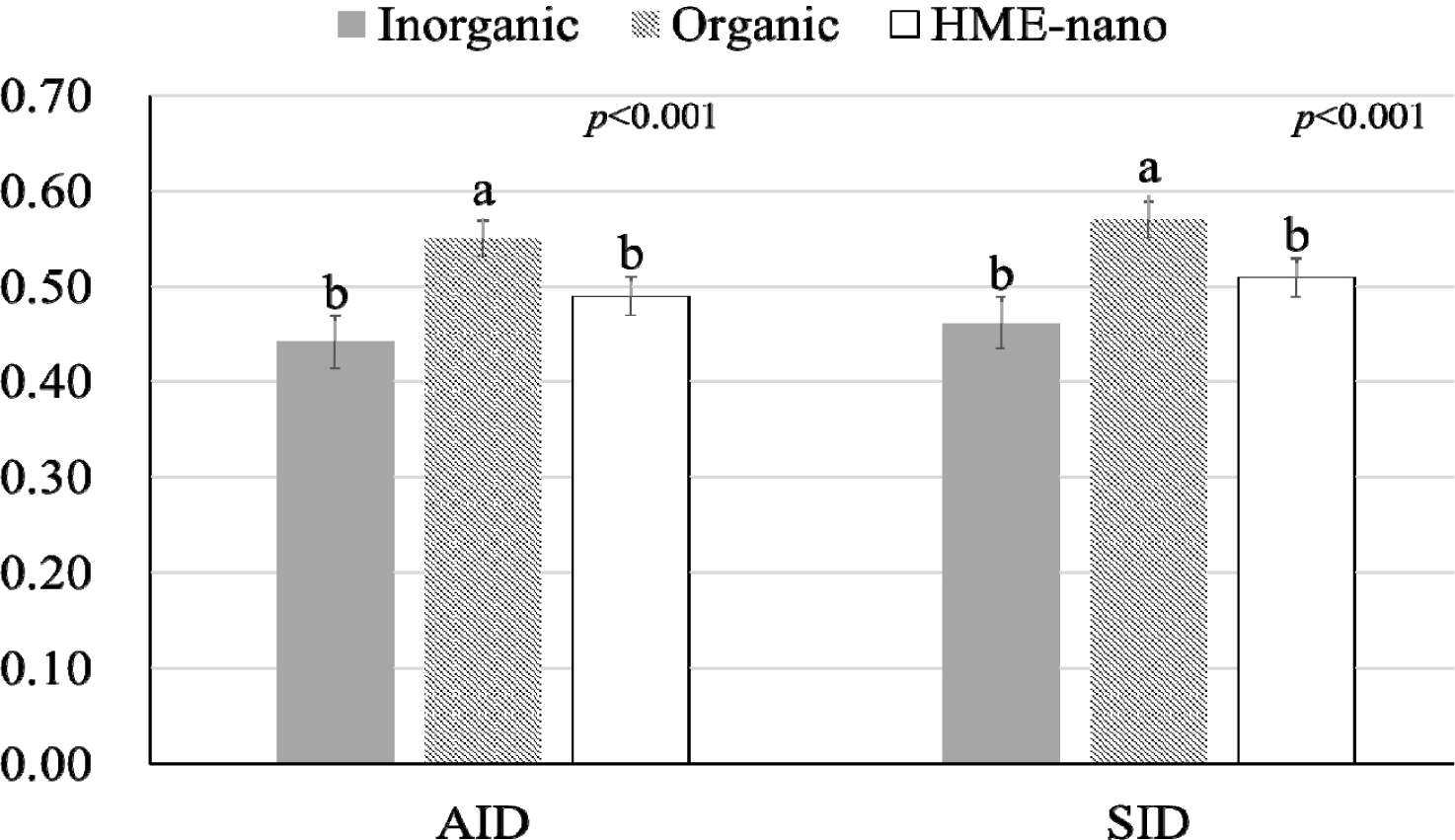
The influence of mineral bioavailability from ileum distal part on different feed ingredients is shown in Figures. There were higher AID and SID values for Cu in corn than in SBM, CGM, and fish meal (p < 0.01; Fig. 1). The AID of Fe was greater in broiler chickens fed corn or fish meal; however, the highest SID of Fe was observed in chickens fed fish meal (p < 0.01; Fig. 2). Moreover, the SID of Fe was higher in chickens fed corn than in those fed SBM or CGM. The AID and SID of Zn in the CGM treatment group were higher than those in the SBM treatment group (Fig. 3). The AID of Mn was higher in broiler chickens fed fish meal compared with those fed CGM and SBM; however, the highest SID of Mn was observed in chickens fed CGM (p < 0.01; Fig. 4). The highest AID and SID of Se were observed in chickens fed fish meal compared to those fed SBM, CGM, and fish meal (p < 0.01; Fig. 5). Moreover, the SID of Se was higher in chickens fed CGM than in those fed SBM or corn. The effects of Cu, Fe, Zn, Mn, and Se source bioavailability are presented in Fig. 6, 7, 8, 9, and 10; respectively. The AID and SID of minerals were higher (p < 0.01) in the nano- and organic forms than in the inorganic form.
DISCUSSION
Cu plays an important role in the growth performance of chickens and needs to be supplemented in the diet of broiler chickens [15–17]. The higher AID and SID availability of Cu in corn than in SBM could be linked to the chemical forms of Cu present in these feed ingredients. CuSO4 is used as Cu source in this study. Moreover, corn contains lower levels of Cu-binding antinutritional factors, such as phytates or oxalates [18,19], which can reduce Cu absorption. Phytates are naturally occurring compounds in plant-based feed ingredients, including SBM [19]. They have strong affinities for minerals such as Cu [3]. When Cu forms complexes with phytates, it becomes less soluble in the gastrointestinal tract of chickens [17]. The reduced presence of phytate in corn results in reduced interference with Cu absorption [3]. As a result, more of the Cu present in corn is available for uptake by chicken intestinal cells. Oxalates are another group of anti-nutritional factors that bind Cu [3,18]. These compounds can form insoluble complexes with Cu, reducing its absorption in the digestive system [18]. In general, corn has a lower concentration of oxalates than SBM [18], contributing to its higher Cu bioavailability.
The increased AID and SID of Fe and Se in fish meal compared with those in CGM and SBM may be due to differences in the chemical forms and nutritional interactions of these ingredients. Fish meal contains Fe and Se in more soluble forms or organically complex minerals, making them more accessible for absorption [1]. Organic complexes provide favorable chemical environments for absorption except phytate [5]. Inorganic minerals such as Fe and Se can form complexes with organic molecules such as amino acids or peptides in fish [1]. These complexes are organically bound or chelated minerals, which increase bioavailability because the surrounding organic molecules can shield the minerals from interactions with other dietary components that might inhibit absorption. This protective effect can enhance mineral absorption by reducing the likelihood of the formation of insoluble complexes with antagonistic compounds [20]. Additionally, organic complexes can serve as specific transport mechanisms that facilitate mineral uptake across the intestinal epithelium [2]. For example, amino acid complexes can be absorbed by amino acid transporters in the gut [8]. Additionally, the presence of free amino acids in fish meal may further enhance mineral solubility and absorption. Amino acids form stable complexes with minerals, thereby improving their transport through the intestinal lining [2,5]. The amino acid profile of fish meal, which is rich in sulfur-containing amino acids such as methionine and cysteine, enhances Se uptake owing to the formation of seleno-amino acids [4], which are more efficiently absorbed.
The lower Zn content in CGM (5.36 mg/kg) than in SBM (14.42 mg/kg) is an important factor to be considered. Zn is absorbed in the small intestine through specific transport systems located on the surface of enterocytes [2,11,21]. These transporters are shared among different minerals, and competition for binding sites can occur when multiple minerals are present [8,9]. When the dietary Zn content is lower in the CGM, there may be less competition for these transporters and receptors. With fewer competing minerals, Zn has a greater opportunity to efficiently bind to available transporters and be taken up by enterocytes [8]. Moreover, the body regulates Zn homeostasis to maintain stable internal mineral concentrations [9]. When dietary Zn intake is low, the body enhances the efficiency of Zn absorption in the intestines to meet physiological needs [9]. Therefore, the body reduces the absorption rate to prevent excessive Zn accumulation. This regulatory mechanism helps maintain the Zn balance.
The substantial difference in Mn content between the meal (3.08 mg/kg) and SBM (13.24 mg/kg) was a critical factor. Mn is absorbed by the small intestine through specific transport systems and receptors. In the small intestine, minerals such as Mn are taken up by enterocytes through specific transporters and receptors [1,22]. These transporters can be shared among various minerals, and competition for binding sites can occur when multiple minerals are present [2,8]. In the case of lower dietary Mn levels, there may be less competition for these transporters and receptors. Reduced competition provides Mn with a greater opportunity to efficiently bind to available transporters and be absorbed by the enterocytes. Moreover, the body has mechanisms that regulate Mn homeostasis to ensure that the internal concentration of Mn remains stable [1,2]. When dietary Mn intake is low, the body can enhance the efficiency of Mn absorption in the intestine to meet its physiological needs. The bioavailability of Mn in fish meal may be higher because of reduced competition for absorption sites. This, in turn, could have resulted in higher AID and SID values for Mn in fish meal than in the SBM.
Organic minerals are typically chemically bound to organic molecules such as amino acids or peptides [4,5]. These organic complexes often mimic forms found in natural feeds. These forms are often bioavailable and are easier for the digestive system to absorb [2,5]. In contrast, nano-minerals refer to minerals that have been reduced to very small particle sizes, often at the nanoscale [6,15,21]. Nano-minerals exhibit unique properties, including increased surface area and improved solubility [10,11]. This enhanced solubility may make the minerals more suitable for absorption. Although the nano- and organic minerals possess different modes of absorption, the fact that there were no significant differences between the organic and nano-minerals suggests that a lower particle size and increased surface area can comparably increase absorption. This result indicates that the nano-minerals in our study were engineered to be highly soluble and effectively absorbed, similar to their organic forms. In some cases, nano-minerals can be tailored to have properties akin to those of organic complexes, enabling them to compete with organic forms in terms of bioavailability. Previous publications have shown a higher absorption of nano-Cu [15,16], nano-Fe [7], nano-Zn [11,21], nano-Mn [22], and nano-Se [23] in the HME form compared with the common inorganic form. Therefore, our study highlights that the organic and nano-forms are superior to the inorganic forms in terms of absorption and that both organic and nano-forms are equally efficient in enhancing bioavailability.
CONCLUSION
The results of this study showed that fish meal had higher bioavailability of Fe, Mn, and Se than CGM and SBM. Corn exhibited the highest Cu bioavailability. The minerals in nano-and organic forms showed higher bioavailability than the common inorganic source, and nano-minerals showed similar bioavailability to the organic form.