INTRODUCTION
The water buffalo (Bubalus bubalis) population in Europe descends from Asian wild buffalo, and the breed is known as the Mediterranean water buffalo. The largest buffalo population in Europe is in Italy, followed by Romania and Bulgaria, and buffalo rearing is on the rise in Southern Europe (Food and Agriculture Organization of the United Nations [FAOSTAT], https://www.fao.org/faostat/en/#home). Buffalo milk contains more proteins, fat, and minerals than cow milk [1,2]. Increasing awareness of buffalo milk’s health benefits consequently has led to the rise in buffalo breeding worldwide and the manufacturing of products from buffalo milk.
Caseins are major milk proteins, accounting for almost 80% of proteins in buffalo milk. There are four types of caseins: αS1, αS2, β, and κ casein, regulated by genes in the same cluster on chromosome 7: CSN1S1, CSN1S2, CSN2, and CSN3, respectively. Casein gene variants can affect milk yield, composition, and processing characteristics [3]. As αS1 casein is the most abundant of all milk proteins, it was the first to be investigated at the protein and gene levels, and associated with milk quality traits. CSN1S1 variant 628C>T, located in exon 17, leads to amino acid change, Ser178 Leu, and this gives αS1 casein variants A and B, respectively. Variant B has shown a significant effect on milk protein percentage [4]. In the buffalo population of Romania, a variation of the αS1 casein B variant has been discovered. The 472G>C substitution in the B variant led to the skipping of exon 6 and the synthesis of a protein lacking eight amino acids [5]. However, the effect of this variant on milk quality traits was not investigated. κ casein has been associated with the stabilization and size of casein micelles [6], traits important for cheese production. It has been established that CSN3 exon 4 codes for most of the mature κ casein. The most significant variant, 467C>T, which leads to an amino acid change, has been associated with a higher percentage of caseins, proteins and fat in cattle milk [7].
In Serbia, buffaloes are reared in small households mainly for local use of milk and butter. Even though the importance of buffalo conservation has been recognized and an in situ program was established nearly twenty years ago [8], there is still a need for selection and refinement. The genetic background of the buffalo population in Serbia has never been investigated, and this is necessary prior to estimating the potential for selection in the Serbian buffalo population. As milk and dairy products are primarily in use, this study aimed to assess the association of casein CSN1S1 and CSN3 gene variants, solely and in haplotype, with milk quality traits in the water buffalo population in Serbia.
MATERIALS AND METHODS
The study involved 130 animals raised in the Raška region (Serbia) within the Pešter area, specifically in Novi Pazar and Tutin municipalities. In the Pešter area, domestic buffalo are extensively reared on pastures, representing an in situ form of conservation. They are primarily raised for milk and butter production, with less emphasis on meat production. During winter, the animals are kept in well-insulated barns as domestic buffalo are not tolerant to cold temperatures. They are taken to pasture in spring and remain there throughout the growing season. Animals were carefully chosen according to the Central Herd Register (Republic of Serbia), so there was no direct kinship between individuals.
The Ministry of Agriculture, Forestry and Water Management of the Republic of Serbia issued an ethical permit for the use of animals for research implementation, number 323-07-10974/2022-05. All rules related to animal welfare were followed.
Blood samples for genetic analyses were collected from the jugular vein (v. jugularis) into 3 mL ethylene-diamine-tetraacetic acid (EDTA)-coated vacuum tubes. The samples were labeled and stored at +4°C until transportation to the laboratory, where they were then preserved at –20°C. DNA was extracted from blood samples of 130 animals using the phenol-chloroform extraction method. CSN1S1 (Gene ID: 102396531) 472G>C and CSN3 (Gene ID: 102395364) 445G>A, 467C>T, 471C>T and 516A>C genotypes were determined using sequencing analysis on an ABI PRISM 3130 Genetic Analyzer (Applied Biosystems). BigDye™ Terminator v3.1 Cycle Sequencing Kit was applied to terminate the polymerase chain reaction (PCR) reaction and for sequencing purposes following the manufacturer’s protocol (Applied Biosystems). Primers I5CZS1-F: 5’ -ACT TAG CAA GGA GAT AAT GCA AGA A-3’ and E7BCZS1-R: 5’ - CTC AGT TGA TTC ACT CCC AAC ATC-3’ were used for amplifying the genomic region from intron 5 to exon 7 of CSN1S1 [5]. Primers 5’ -CGC TGT GAG AAA GAG GAA AGA TTC-3’ and 5’ -AGA TTC AAG GAG TAT ACC AAT TGT TG-3’ were used for amplifying exon 4 of CSN3 [9]. Forward primers were utilized for the sequencing analysis. Detection of genotypes was conducted using Sequencing Analysis Software V4.0 (Applied Biosystems). Genotyping succeeded in 128 samples for the CSN1S1 variant and 125 for the CSN3 variants. A quarter of the samples were genotyped for CSN1S1 472G>C by the PCR-restriction fragment length polymorphism (RFLP) method. The PCR product was digested with TaaI restriction enzyme, and the products of digestion were visualized on an 8% polyacrylamide gel stained with silver nitrate. Genotypes were consistent with the results of sequencing.
Milk samples were collected from a subgroup of 42 animals in the same lactation stages three times at equal intervals. Immediately after sampling, the milk was cooled, and no preservatives were added to the samples. Milk was sampled during winter when the animals were housed in barns. The composition of raw buffalo milk samples was determined by the following methods: titratable acidity according to the Soxhlet-Henkel method [10], total solids by the standard drying method at 102 ± 2°C [11]; fat contents according to the Gerber method [12]; nitrogen and casein nitrogen contents by the Kjeldahl method [13], while the protein and casein contents were calculated as the nitrogen contents and casein nitrogen contents multiplied by 6.38.
Deviations from Hardy-Weinberg equilibrium were assessed by the chi-square (χ2) test. Values of continuous variables are presented as mean ± SD. Casein correlations with other investigated milk quality traits were analyzed with the Spearman test and presented as Spearman R-value. The t-test was used to compare the values of continuous variables with a normal distribution within two groups. The nonparametric Mann–Whitney U test was used to compare the values of continuous variables with a skewed distribution within two groups. The strength of the association between CSN1S1 and CSN3 genotypes and milk components was assessed using a multiple linear regression model and presented as β coefficient. For the regression analysis we have applied linear model as follows:
where:
1. Y is the dependent variable (e.g., casein level in milk).
2. β0 is the intercept of the regression line.
3. β1 is the coefficient representing the effect of the first genetic variant (X1).
4. β2 is the coefficient representing the effect of the second genetic variant (X2).
5. ϵ is the error term, which accounts for the variation in Y not explained by the linear relationship with X1 and X2.
The proportion of the variance in the casein and protein level that can be explained by the CSN1S1 gene variant is presented as the R2 from the regression model. Homogeneity of Variance was confirmed by the Levene’s Test. The p-value < 0.05 was considered statistically significant. Statistical analyses were performed using the Statistica Version 8 software package [14].
Haplotype analysis was performed by the publicly available Thesias software (www.genecanvas.org). Thesias applies the stochastic-expectation–maximization (EM) algorithm to estimate haplotype frequencies and their associated effects on the phenotype of interest [15,16]. For haplotype-phenotype association it uses the likelihood ratio test. Thesias assumes the additivity of the haplotype effects. The software then fits the following linear model to estimate the effects of haplotypes on the continuous trait. The Thesisas software was also used to estimate the linkage disequilibrium (LD) parameters within the studied groups and to calculate the percentage of variance explained by the haplotypes. According to the Thesias software manual, CSN1S1 and CSN3 haplotypes’ effect on milk quality trait levels are given as mean value for one dose of each haplotype along with 95% CI compared to the reference, most frequent haplotype. A p-value < 0.008 was considered statistically significant because we applied Bonferroni correction for six milk quality traits being assessed. Statistical power of the study was calculated by the on-line Post-hoc Power Calculator (https://clincalc.com/stats/Power.aspx) [17].
RESULTS
Variants, CSN1S1 472G>C (Supplementary Fig. S1) and CSN3 467C>T were in Hardy–Weinberg equilibrium in the Serbian Buffalo population. The genotype distribution and allele frequencies are presented in Table 1. Three variants were additionally investigated, two of them, CSN3 445G>A and CSN3 516A>C, were not polymorphic in the investigated Serbian Buffalo population, meaning that only wild-type alleles G for CSN3 445G>A and allele A for CSN3 516A>C, were present. And, the third variant CSN3 471C>T was in complete LD with variant CSN3 467C>T (Supplementary Fig. S2).
Table 2 presents the composition of milk for the studied subgroup of 42 buffaloes and correlations between milk casein percentage and the other five milk quality traits. Caseins were significantly positively correlated with protein, acidity SH°, total solids, and solids-not-fat, and were not significantly correlated with milk fat percentage.
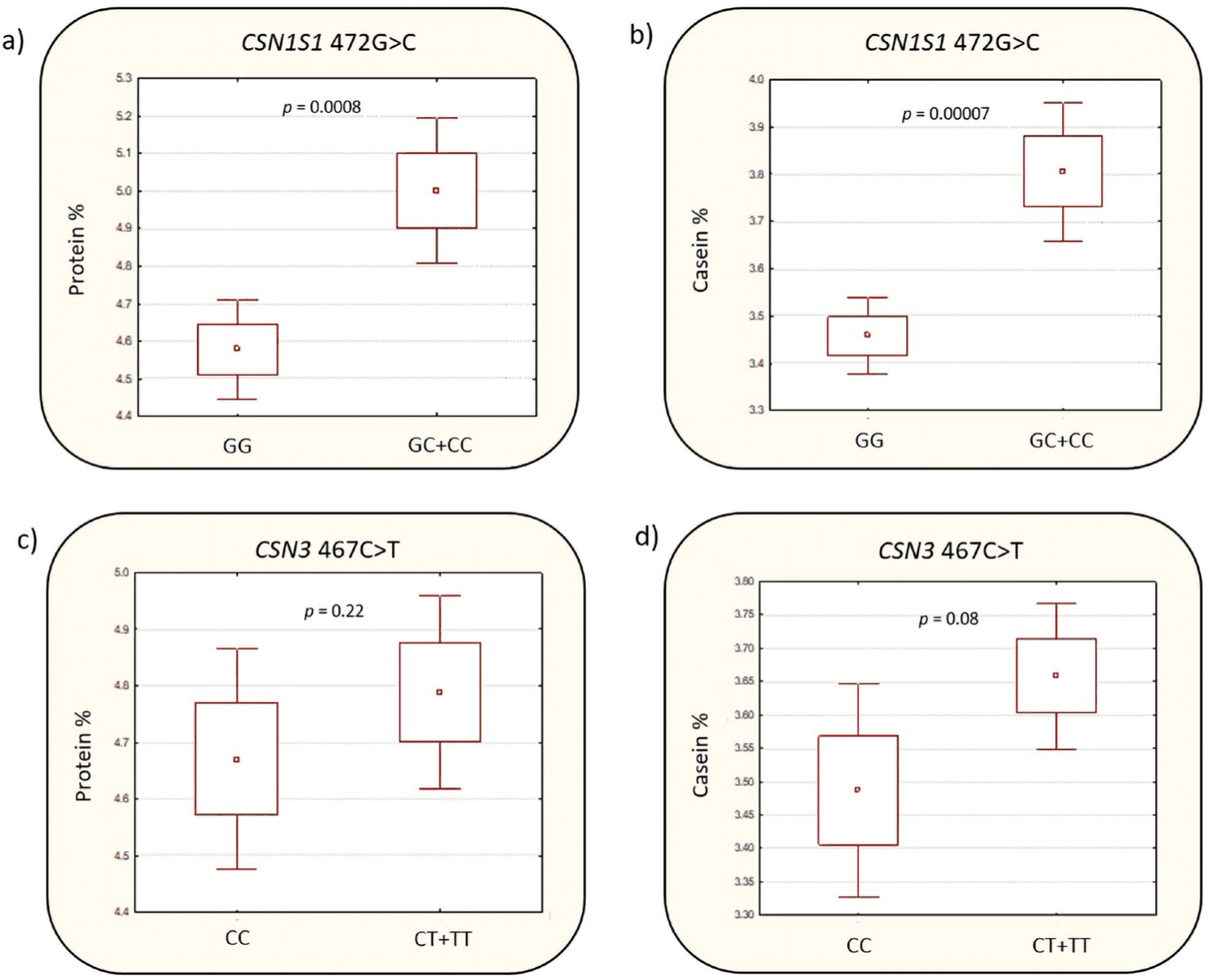
Fig. 1 shows individual associations of variants CSN1S1 472G>C and CSN3 467C>T with protein and casein levels in milk. We observed that minor allele C (CSN1S1 472G>C), by the dominant model of inheritance, GG vs. GC+CC, was associated with higher protein levels in milk, (p = 0.0008, t-test) and higher casein levels, (p = 0.00007, t-test). The study power for this association was 90% for α = 0.05. Proportion of variance in caseins explained by CSN1S 472G>C was 33%, and for proteins it was 25%, by the regression model. Variant CSN3 467C>T was not associated with protein and casein levels in milk by the dominant model of inheritance, CC vs. CT+TT. However, by the multiple linear regression model, we found significant effects of CSN1S1 472G>C (β = 0.62, p = 0.000013) and CSN3 467C>T (β = 0.32, p = 0.012) on milk casein levels. Substitution effect for variant CSN1S1 472G>C, individually, is β = 0.58 for caseins and β = 0.50 for proteins.
As CSN1S1 and CSN3 are located on chromosome 7 but are not in a strong LD (r2 = 0.17, D’ = –1), there was a significant rationale to analyze them as haplotype. In Table 3 are presented haplotype frequencies of CSN1S1 472G>C and CSN3 467C>T and their effects on milk quality traits. The Thesias software set the 1G3C haplotype as a reference because it was the most frequent haplotype in the investigated buffalo population. Compared with the reference haplotype 1G3C (mean casein level [95% CI] = 3.34 [3.18–3.50], haplotypes 1G3T and 1C3C had significantly higher casein levels in milk (1G3T - mean casein level [95% CI] = 3.68 [3.48–3.88], p = 0.02 and 1C3C - mean casein level [95% CI] = 3.90 [3.72–4.06], p = 0.00002). In addition, haplotype 1C3C had significantly higher protein levels (mean protein level [95% CI] = 5.14 [4.88–5.40], p = 0.0004) in comparison with the reference haplotype 1G3C (mean casein level [95% CI] = 4.52 [4.32–4.72]). To provide reliable results, we set stringent significance cut-off values by applying the Bonferroni correction (p < 0.008), and consequently, we did not consider the association of 1G3T haplotype with casein level significant. The study power for haplotype effect on casein levels was 85% for α = 0.008, and for haplotype effect on protein levels was 65% at the same stringent significance level of 0.008. The percentage of variance explained by these haplotypes was 40% for caseins and 31% for proteins.
DISCUSSION
The main buffalo species in Europe is the water buffalo, with the Mediterranean water buffalo as the predominant breed. There are genetic variants among geographic regions, and their detection is essential for the authentication of protected denomination of origin (PDO) products. These tests are often based on analysis of the major milk protein genes. Casein genes have a great impact on milk quality, as expected because caseins are the most abundant proteins in milk. Also, casein genes are of interest in evolutionary studies, as well as in selection and refinement [18]. According to the Food and Agriculture Organization of the United Nations (FAO)/Domestic Animal Diversity Information System (DAD-IS); https://www.fao.org/dad-is/browse-by-country-and-species/en/), in Serbia, the buffalo population is increasing, but it is still marked as endangered. It is crucial to genetically characterize the local buffalo population for future selection and breeding. Knowledge of the genetic background and careful selection are essential to avoid inbreeding and to maintain genetic diversity in the population [19]. Inbreeding has various adverse effects on milk production and quality traits due to the small effective size of the population, decreased response to selection, and reduced animal performance. Significant inbreeding has already been noticed in buffalo populations [20], and the genetic potential of never-selected buffalo populations might be useful for reintroduction purposes [21]. Milk protein and fat percentages in the investigated Serbian buffalo population were comparable with other buffalo populations [3,22,23], though different from the milk quality reported in a previous study on buffalos from the Pešter region [24], which reported much lower fat percentages in milk.
In this study, we have analyzed the CSN1S1 gene variant, which was detected in the neighboring Romanian buffalo population but has not been found in any other buffalo population [5]. CSN1S1 472G>C is an intronic variant that leads to skipping exon six and produces a protein lacking eight amino acids. We detected this variant in the Serbian buffalo population of 128 animals with a frequency of 0.23 for the minor C allele, while in the Romanian buffalo population of 160 animals, it was 0.18. We have found an association between the variant CSN1S1 472G>C and the milk casein and protein levels. Even though Balteanu et al. implied that the 472G>C variant causes a defective protein, functional studies are needed to support that premise. Our results suggest that milk from animals with the minor C allele has more caseins and proteins. The association of variant CSN1S1 472G>C with milk quality traits has not been previously analyzed, so our results are the first regarding this variant’s effect on milk composition. It would be of interest to explore the presence of this variant in other local neighboring populations so we could define our buffalo population in terms of milk protein-coding genes compared with those of neighboring populations. A limitation of our study was that we measured the total casein percentage in milk samples and not the percentage of specific casein types, which would have given us information on αS1 casein presence in milk samples. As shown in cattle [7], variants in casein genes affect the concentration of the specific caseins in milk. T allele of gene variant CSN3 467C>T was associated with a lower concentration of αS1 casein and a higher concentration of αS2 and κ casein in cow milk.
Variant CSN3 467C>T has been explored by others in buffalo breeds, but in smaller sample sizes and mainly not in association with milk quality traits [3,9,25–28]. Fan et al. performed an extensive analysis of CSN3 gene variants. It has shown that in Chinese water buffalo population frequencies of minor alleles were found to be less than 0.1 for investigated variants (CSN3 445G>A, 467C>T 471C>T, and 516A>C) [9]. This indicates that the Chinese water buffalo population is very different from other water buffalo populations around the world due to migration and crossbreeding events. Since, variant CSN3 467C>T was among these investigated variants, it gave us the reason to assume that other variants investigated in that study could be polymorphic in Serbian buffalo population. Nevertheless, CSN3 variants 445G>A and 516A>C were not polymorphic in our buffalo population and CSN3 471C>T was in complete LD with variant 467C>T. Variant 471C>T is silent and it does not lead to an amino acid change, while CSN3 467C>T changes the amino acid threonine to isoleucine and has a functional effect on κ casein protein. This variant is the only CSN3 variant shared between buffalo and cattle, while other variants found in buffalo CSN3 gene do not exist in cattle, and vice versa [9]. Allele T of the variant CSN3 467C>T has been associated with higher casein, protein, and fat content in cow milk [7]. Our study analyzed six milk quality traits, and the minor allele T showed a significant effect on a casein level in milk. Of previously mentioned studies, just one addressed milk quality traits and associated allele T with higher protein and casein levels in buffalo milk [26].
We have analyzed CSN1S1 472G>C and CSN3 467C>T haplotype effects on milk quality traits. Haplotype analysis can provide insight into the true effect of variants inherited together in a population. The most abundant haplotype in the investigated Serbian buffalo population, with a frequency of 0.40, was haplotype 1G3C, inferred from wild-type alleles of both variants. By comparison, haplotype 1C3C consisted of CSN1S1 472G>C minor allele C and CSN3 467C>T wild type allele C, had higher levels of caseins and proteins in buffalo milk. So far, only two variants in casein genes have been analyzed in haplotype CSN1S1 628C>T and CSN3 467C>T, and mainly with regard to milk yield and coagulation properties [3,26]. Our individual analysis showed that allele T of gene variant CSN3 467C>T was associated with higher levels of caseins in milk, and we confirmed this in haplotype analysis, showing that haplotype 1G3T had a higher level of caseins than haplotype 1G3C. Despite this, the haplotype 1C3C had the largest increase in milk casein levels. Based on individual variant analysis, it would be expected that the 1C3T haplotype has the greatest effect on casein, but this haplotype is so rare that it was not even detected in our buffalo population.
In conclusion, we have shown that genetic variants CSN1S1 472G>C and CSN3 467C>T have a significant impact on casein levels in buffalo milk. CSN1S1 minor allele C was associated with higher levels of caseins and proteins in milk. In addition, haplotype analysis of these genetic variants provides greater validity and reliability in association with casein levels compared with individual analysis. Our results contribute to knowledge of the buffalo genome by characterizing the buffalo population in Serbia, which has not been performed to date, and provide potentially valuable perspectives for applications in improving milk quality.