INTRODUCTION
Egg protein has been identified as being highly digestible and a great supply of essential amino acids, with the highest achievable protein digestibility-corrected amino acid score [1], which is driving increased global demand for egg consumption. The growing worldwide consumption of eggs has driven the expansion of intensive poultry production operations, which can unfortunately result in elevated environmental stress factors. Environmental stressors including temperature, humidity, and stocking density are the main factors affecting animal welfare, health, and productivity [2]. Geng et al. [3] reported that stocking density in rearing spaces has become one of the most important environmental and management factors for modern intensive animal husbandry. Nevertheless, in order to increase total egg production per housing unit, many egg producers endeavor to decrease the payback period and increase their net income by increasing the number of hens per cage at maximum capacity [4].
With increasing stocking density (342 to 690 cm2/hen), egg performance and metabolizable energy (ME) efficiency for egg production decreased significantly in hens kept at a stocking density of 342 cm2 per hen [5]. A dense environment also was associated with detrimental impacts including decreased egg production and egg mass [6], decreased laying rate, and increased levels of noxious gas emissions from the litter [7]. Wang et al. [8] found that high stocking density reduced the laying rate and eggshell quality in laying hens. Physiologically, birds kept at high stocking density may be more susceptible to oxidative stress [9,10]. Incharoen et al. [11] noted that nutritional modification might be a key factor to help ameliorate stress from high stocking density in laying hens. Thus, addition of specific antioxidants to the diet could be one efficient approach to alleviate the negative impact of stress [12].
Gamma-oryzanol (GO) in the rice bran layer has been identified as a potent natural antioxidant due to its capacity to prevent lipid peroxidation and the resulting oxidative stress [13]. It contains a mixture of ferulic acid esters and phytosterols (sterols and triterpenic alcohols) [14,15]. Antioxidant components of GO such as 24-methylenecycloartanyl ferulate, cycloartenyl ferulate, campesteryl ferulate, and β-sitosteryl ferulate are able to inhibit lipid peroxidation and free radical production, and scavenge the free radicals from the body [16]. Ferulic acid not only scavenges free radicals, but also enhances the activity of these enzymes. Additionally, it inhibits enzymes that produce free radicals [17]. The structure of GO components is similar to that of cholesterol and can reduce oxidative stress and maintain the functionality of cells [18].
As a lipid-soluble nutrient, vitamin E takes on a vital function as a peroxyl radical-scavenging antioxidant that inhibits lipid peroxidation by breaking chain propagation [19]. Natural vitamin E consists of 8 different analogues: α-, β-, γ- and δ-tocopherol; and α-, β-, γ- and δ-tocotrienol [20]. Among these, α-tocopherol has been mainly used as a supplement in livestock feed. Compared with a basal diet without vitamin E supplementation, dietary α-tocopherol acetate (200 to 500 mg/kg) enhanced the antioxidant capacity in hens [21–23]. Zhao et al. [24] reported that dietary natural tocopherol at a dosage of 100 mg/kg enhanced laying performance and tocopherol deposition as well as regulated serum cholesterol concentrations and improved antioxidant status. Although tocopherol is the most accepted analogue for feeding domestic animals, Serbinova et al. [25] reported higher antioxidant activity against lipid peroxidation with tocotrienol than with α-tocopherol in rat liver microsomes. Thus, it appears that vitamin E tocotrienol (VE) has greater potency. Furthermore, tocotrienols possess powerful antioxidant, neuroprotective, anti-cancer, and cholesterol regulatory activities that often differ from the properties of tocopherols [26].
Because of the reported adverse effects of high stocking density on hen health and productivity, we hypothesized that dietary supplementation with GO and VE, individually or in combination, could mitigate these negative impacts. We aimed to assess the effects of these supplements on productivity, egg quality, and immune and health-related mRNA levels in laying hens raised at varying stocking densities.
MATERIALS AND METHODS
Hy-Line Brown layers purchased from a commercial farm in Phitsanulok province were used. The GO (98.0% purity) and VE (60.0 % total tocotrienols and 30.0 % total tocopherols) products were extracted from the rice bran of Oryza sativa Linne (Gramineae) and obtained from Oryza Oil & Fat Chemical. All animals were reared in wire cages in tunnel-ventilated houses equipped with an evaporative cooling system to control the ambient temperature. Throughout the duration of the experiment, the average temperature remained consistent at 28 ± 2°C, accompanied by a relative humidity range of 60%–65%. LED artificial lighting was at a consistent photoperiod (17L:7D). At 54-weeks of age, a total of 120 laying hens with identical body weight (1,920 ± 50 g) and egg uniformity were allocated into 8 groups with 5 replicates per group (3 layers/replicate). A completely randomized experiment with a 2 × 2 × 2 factorial arrangement of treatments was used to evaluate the effect of stocking density (low vs. high) and an addition of 200 ppm GO, 200 ppm VE, or both on productivity, egg quality, and immune- and health-related mRNA abundance. The first 4 groups of hens were kept in wire cages with a low stocking density of 840 cm2/hen. Laying hens in the other 4 treatments were confined to a wire cage with a high stocking density of 420 cm2/hen. During the 54 to 62 weeks of age, all hens had free access to clean drinking water and feed. Diets were formulated in accordance with the nutrient requirement recommendation of NRC [27] (Table 1).
Vitamin-mineral premix provided per kilogram of diet: vitamin A (trans-retinyl acetate), 12,000 IU; vitamin D3 (cholecalciferol), 3,000 IU; vitamin E (allrac-tocopherol-acetate), 12 mg; vitamin K3 (bisulphate menadione complex), 3.6 mg; vitamin B1, 1.4 mg; vitamin B2, 5.4 mg; vitamin B6, 4.2 mg; vitamin B12 (cyanocobalamin), 0.02 mg; nicotinic acid, 9 mg; pantothenic acid, 9 mg; folic acid, 0.6 mg; biotin, 45 mg; choline chloride, 210 mg; selenium, 0.18 mg; cobalt, 0.3 mg; iodine, 1.08 mg; iron, 54 mg; zinc sulfate, 60 mg; manganese oxide, 96 mg; copper sulfate, 12 mg.
The nutrient values were calculated based on the analyzed nutrient values according to NRC [27].
All eggs were carefully collected twice daily (at 6:00 AM and 6:00 PM) from each cage and counted in each replication. We also recorded the weight of each collected egg on a daily basis, while monitoring the remaining feed on a weekly basis. Parameters analyzed included hen-day egg production, average egg weight, average daily feed intake, egg mass, and feed conversion ratio. Additionally, we collected 10 eggs from each group on a weekly basis to assess eggshell breaking strength, eggshell thickness, eggshell ratio, yolk ratio, albumen ratio, albumen height, yolk color and Haugh unit. These parameters were evaluated using a TA-XT2 Plus Analyzer (Stable Micro systems) following the methods described by Likittrakulwong et al. [28].
At 62 weeks of age, blood samples were collected from five hens from each group. They were taken by venipuncture from the wing vein, and blood was saved into collection tubes using a sterile syringe, kept in blood collection tubes, and stored at 4°C in a refrigerator. Blood samples were mixed with an anticoagulant solution (ethylene diamine tetraacetic acid [EDTA]) and then used for mRNA abundance analysis of 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGCR) and heat shock protein 70 (HSP-70) [11]. After blood collection, hens were sacrificed under mild anesthesia. Whole visceral organs were pulled out of the abdomen and placed on a clean aluminum tray. Using sterile equipment, the spleen was removed and cut into a small pieces of 4–5 mm thickness, rapidly frozen in liquid nitrogen, and kept at –80°C until mRNA abundance analysis of interleukin-12 subunit beta (IL-12β) and interferon gamma (IFN-γ). Approximately 30 mg of spleen tissue from each treatment were homogenized with the TissueRuptor homogenizer (Qiagen GmbH, Hilden, Germany) in 350 µL of RLT buffer (RNeasy Mini RNA isolation kit, Qiagen GmbH) and stored at −80°C for RNA extraction.
Total RNA was isolated using the RNeasy Mini RNA isolation kit (Qiagen GmbH,) and eluted in 50 µL RNase-free water. The concentration of total RNA was measured using a nanodrop Quawell UV–VIS Spectrophotometer Q5000 (Quawell Technology). One µg of total RNA from each sample was used for first-strand cDNA synthesis, which was performed using the RevertAidTM first strand cDNA synthesis kit (Fermentas), following the manufacturer’s recommendations. One µL of first-strand cDNA from each sample was used as the template for semi-quantitative reverse transcription polymerase chain reaction (RT-PCR) analysis. PCR amplification was performed using specific primers [29–31] (Table 2). Quantitative Real-time RT-PCR (qPCR) was performed as previously described by Incharoen et al. [11] to measure the levels of HMGCR, HSP70, IL-12β, IFN-γ and beta-actin (internal control) mRNA. The reactions were performed in triplicate in a MyGo Pro real-time PCR instrument (IT-IS Life Science). The relative mRNA abundance was analyzed using MyGoPro qPCR software (IT-IS Life Science). Results of real-time PCR were analyzed by the 2-ΔΔCt method [32]. The mRNA abundance of these genes was normalized to beta-actin.
| Gene | Sequence (5’- 3′) | Annealing temperature (°C) | Product size (bp) | References |
|---|---|---|---|---|
| HMGCR |
F:ATGCATGGCCTTTTTGTGGCCTCTCATCCA R:CTTGAGAAGATTGTGAGGAGACCAGCAATA |
55 | 242 | Beloor et al. [29] |
| HPS70 |
F:AATCTATCATCATGTCTGGCAAAGGGCCGG R:GCGGCCGATGAGACGCTTGGCATCAAAGAT |
58 | 220 | Beloor et al. [29] |
| IL-12β |
F:TGTCTCACCTGCTATTTGCCTTAC R:CATACACATTCTCTCTAAGTTTCCACTGT |
60 | 82 | Brisbin et al. [30] |
| IFN-γ |
F:ACACTGACAAGTCAAAGCCGC R:AGTCGTTCATCGGGAGCTTG |
60 | 129 | Brisbin et al. [30] |
| β-actin |
F:CCACCGCAAATGCTTCTA R:GCCAATCTCGTCTTGTTTTATG |
60 | 96 | Sohn et al. [31] |
Data on egg performance and quality were subjected to analysis of variance using the General Linear Models (GLM) procedures of SPSS version 17.0 (SPSS) according to a 2 × 2 × 2 factorial arrangement of treatments, including stocking density (low vs. high) and an addition of GO (none vs. 200 ppm) or VE (none vs. 200 ppm) as the main effects and the respective interactions. One-way ANOVA was used to determine differences in mRNA abundance. Statistically significant means were compared using Duncan’s Multiple Range Test and a probability level of p < 0.05 was considered significant.
RESULTS
There was a main effect of stocking density (p < 0.01) on average daily feed intake and hen-day egg production of laying hens where compared with laying hen housed at low stocking density, high stocking density resulted in a significantly lower average daily feed intake and hen-day egg production during 54–62 weeks of age (Table 3). Regardless of stocking density and dietary GO, the supplementation of VE increased significantly average egg weight and egg mass (p < 0.05). However, an improved feed conversion ratio and higher egg mass were detected in laying hens fed dietary GO (p < 0.05). A significant interaction of VE by GO supplementation was observed on feed conversion ratio (p < 0.05). This was accompanied by improved feed conversion ratio of laying hens reared under high stocking density. In contrast, no interactions among dietary VE, GO and stocking density were observed on overall performance during the experimental period (p > 0.05).
Animal were reared in different stocking densities: LSD, low stocking density (840 cm2/bird); HSD, high stocking density (420 cm2/bird). They were fed with different diet including CON, a basal diet without supplementation; GO, a basal diet supplemented with 200 ppm gamma-oryzanol; VE, a basal diet supplemented with 200 ppm vitamin E tocotrienols; GO + VE, a basal diet supplemented with 200 ppm gamma-oryzanol + 200 ppm vitamin E tocotrienols.
Except for eggshell breaking strength (p < 0.05), there was no main effect of dietary VE, GO and stocking density on eggshell thickness, eggshell ratio, yolk ratio, albumen ratio, albumen height, yolk color and Haugh unit (Table 4). This was associated with decreased eggshell breaking strength in laying hens reared under high stocking density, while eggshell breaking strength improved in hens fed dietary VE and GO. There were no significant interactions of dietary VE, GO and stocking density on overall egg quality parameters (p > 0.05).
Animal were reared in different stocking densities: LSD, low stocking density (840 cm2/bird); HSD, high stocking density (420 cm2/bird). They were fed with different diet including CON, a basal diet without supplementation; GO, a basal diet supplemented with 200 ppm gamma-oryzanol; VE, a basal diet supplemented with 200 ppm vitamin E tocotrienols; GO + VE, a basal diet supplemented with 200 ppm gamma-oryzanol + 200 ppm vitamin E tocotrienols.
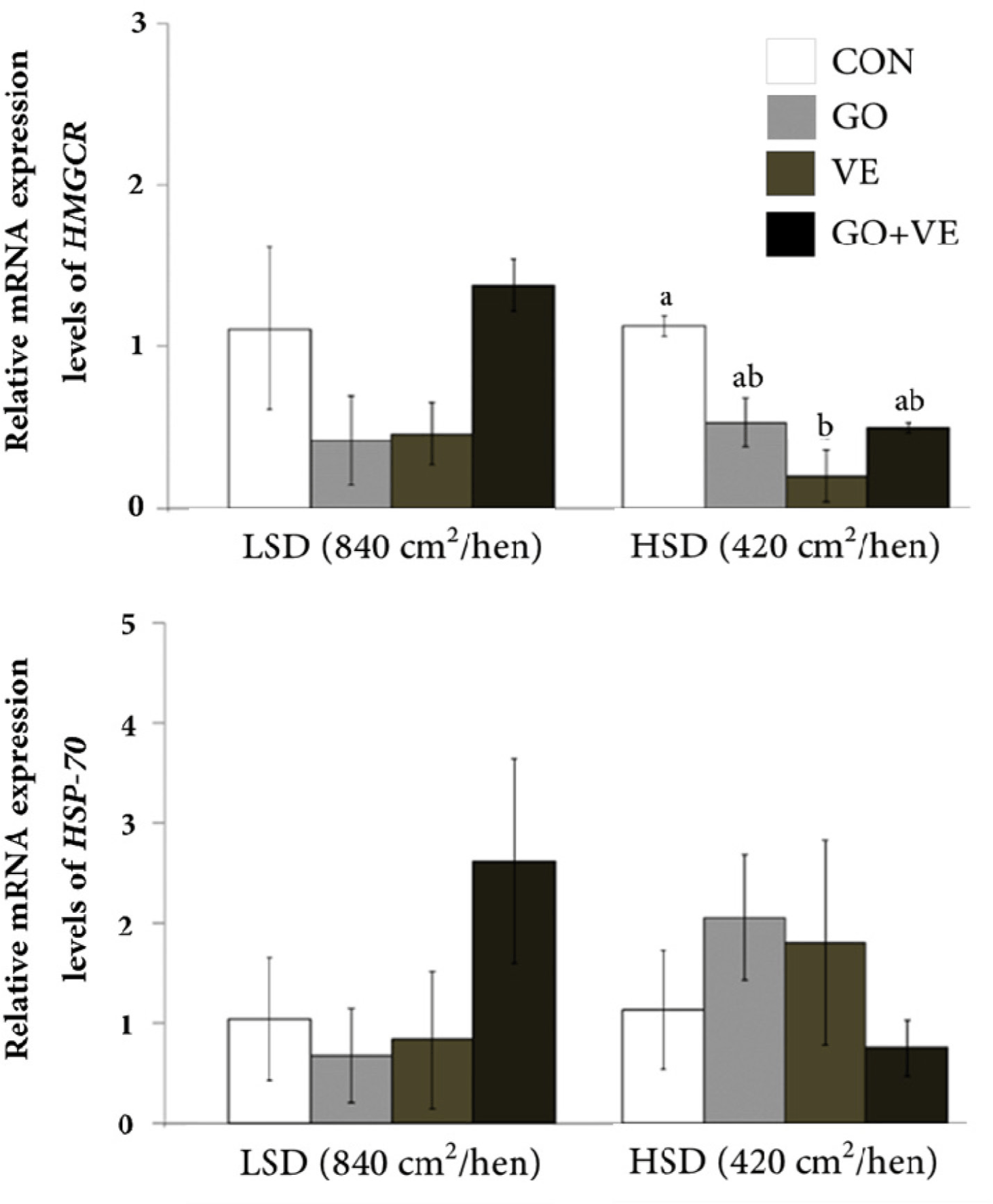
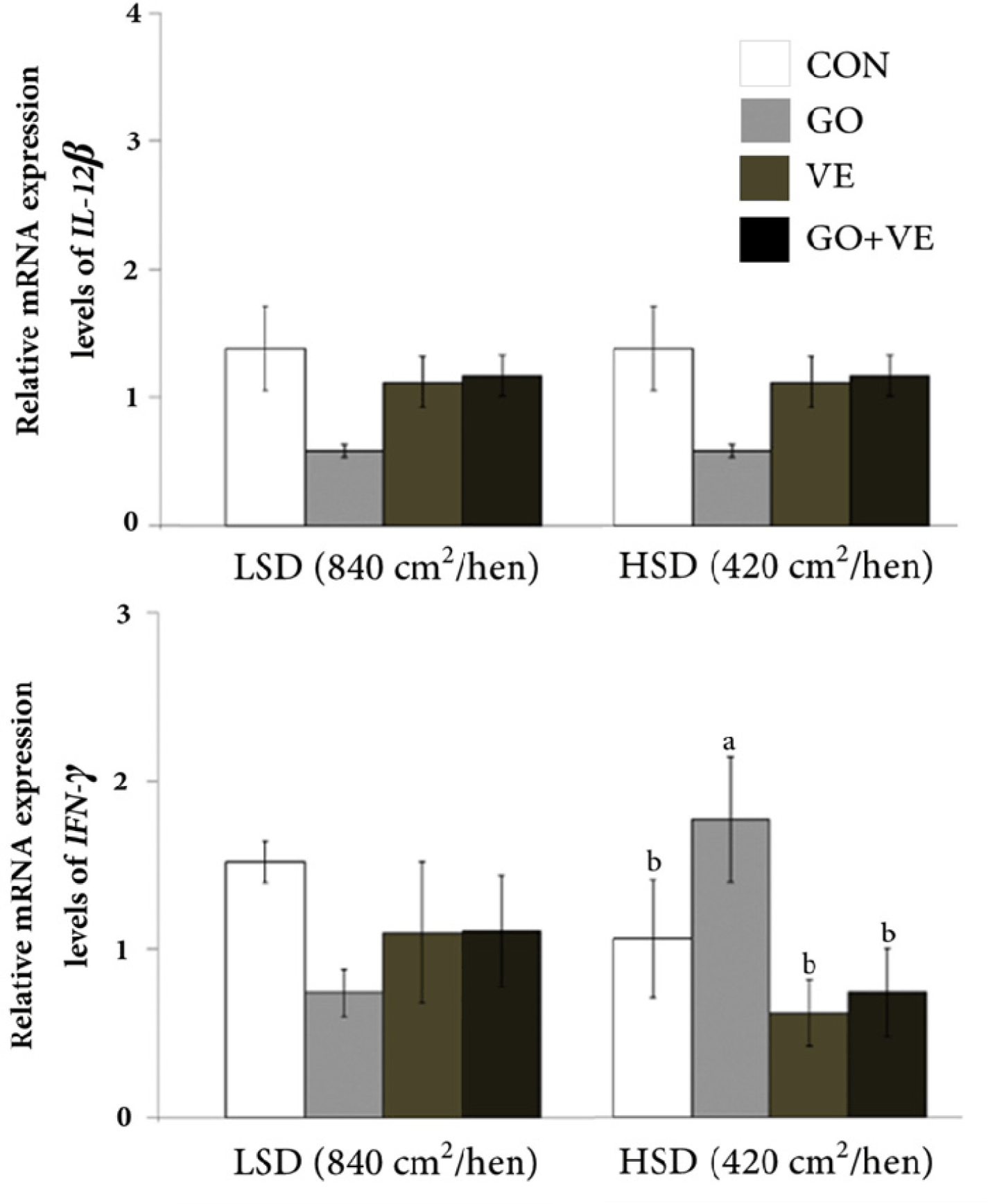
In the low stocking density conditions, the laying hens had minimal differences in mRNA abundance of HMGCR, HSP70, IL-12 β, and IFN- γ genes across all dietary treatments (Figs. 1 and 2). However, laying hens housed at high stocking density had a decrease (p < 0.05) in HMGCR abundance within the VE group compared with the CON group. Among animals kept in high stocking density conditions, those fed dietary GO exhibited the highest level of IFN-γ mRNA abundance relative to other diets. However, there was no difference in abundance of HSP70 and IL-12 β in laying hens reared under high stocking density regardless of diet.
DISCUSSION
The reduction in average daily feed intake, hen-day egg production, and eggshell breaking strength observed in laying hens raised under high stocking density agreed with the recent study from Incharoen et al. [11] in which heat stress-induced hens kept in high stocking density displayed decreased egg performance and eggshell-breaking strength. Similarly, the laying hens kept in low stocking density had considerably higher feed intake and ME intake than those reared in high stocking density [5]. Anderson et al. [6] also reported that hens housed at a high stocking density had reduced egg mass and hen-day egg production. There are also some detrimental effects of increasing the stocking density on gas emissions, litter moisture content, and laying efficiency [7]. In broilers, there is evidence confirming that increasing stocking density decreased feed intake, body weight, weight gain, and feed conversion ratio [33,34]. Goo et al. [35] also reported that broilers reared under heat stress and high stocking density decreased performance with a negative impact on breast meat quality. During the starter period, studies have observed that high stocking density is associated with a reduction in feed intake and weight gain in White Pekin ducks [36] and geese [37]. Furthermore, research studies have provided scientific evidence that housing birds in a densely stocked environment can lead to elevated ambient temperatures surrounding the birds and lowering body heat dissipation resulting in heat stress conditions [34,38]. Thus, this overwhelming evidence underscores that monitoring environmental factors and adjusting nutritional management practices accordingly is crucial to minimize harmful outcomes on health and welfare.
To our knowledge, there is no published research on the potential benefits of incorporating GO and VE, either individually or in combination, into poultry diets with regard to reducing the detrimental effects of oxidative stress caused by high stocking density. However, Minatel et al. [13] demonstrated that GO in the rice bran layer is a potent natural antioxidant due to its capacity to prevent lipid peroxidation and the resulting oxidative stress. In addition, López-Revuelta et al. [18] noted that the structure of GO components is analogous to that of cholesterol, meaning that it can help reduce oxidative stress and support the normal functioning of cells. Previous studies have also reported that GO has the potential to positively affect the immune system, lipid levels in blood, antioxidant capabilities, and better efficiency for the animal to avoid heat stress [39–41]. Kang and Kim [42] noted that dietary rice bran oil containing GO as natural antioxidant improved feed conversion ratio of broiler chickens during 0 to 35 days of age. Broiler chickens consuming dietary rice bran oil with high levels of GO (3.58 g/100 g oil) had a significantly lower feed conversion ratio resulting in improved growth performance [43]. According to our results, feed conversion ratio and egg mass were significantly enhanced in laying hens fed dietary GO. Although the mechanism explaining this phenomenon could not be discerned, the inclusion of oryzanol in the laying diet was advantageous.
Tocopherol is the generic form of vitamin E used in feed because of its effectiveness as an antioxidant and inhibitor of lipid peroxidation by breaking chain propagation [19]. Previous research provided evidence that hens fed a diet containing α-tocopherol acetate had stronger antioxidant capacity than control-fed birds [21–23]. Recently, Zhao et al. [24] reported that dietary tocopherol content (100 mg/kg diet) increased egg-laying performance and tocopherol deposition as well as regulated serum cholesterol concentration and improved antioxidant status. Furthermore, vitamin E supplementation increased egg production and quality while also providing health advantages to laying hens fed a diet rich in corn dried distillers grains with solubles [44]. In the current study, a significant improvement in average egg weight and egg mass was detected in laying hens receiving dietary VE. This suggested that vitamin E’s advantageous properties were due to its ability to facilitate the release of vitellogenin, thus, stimulating egg formation [24,45]. The lack of significant effect of VE on increased egg-laying performance was likely due to the fact supplementation of VE exceeded the nutritional requirements for egg production of the laying hens. As a result, VE primarily had antioxidant effects in the heat-stressed hens under increasing stocking density.
In this study, the eggshell breaking strength significantly decreased in the hens exposed to high stocking density during the whole experimental period. These results are consistent with those of some animal nutritionists, who reported that the eggshell strength decreased significantly in heat-stressed laying hens [46,47]. The present findings revealed that the decrease in eggshell quality was likely due to a significant reduction in daily feed intake in laying hens reared under high stocking density, which reduces the amount of essential minerals such as Ca, Mg, and P available for egg formation. In addition, decreased productivity and poor egg quality were observed in hens raised under high stocking density, possibly due to reduced digestibility caused by heat stress [48]. However, compared with the unsupplemented group, eggshell breaking strength increased significantly in hens fed dietary GO and VE.
There is some evidence suggesting that antioxidants delivered through supplementation in the diet can minimize oxidative stress [49,50] leading to enhanced growth and feed efficiency, and optimizing nutrient utilization. Others reported that high dietary concentrations of GO, VE, and other bioactive components in rice bran oil improved growth performance of broiler chickens [42,51]. The present data demonstrating a significant interaction of GO and VE were related to improved feed conversion ratio of laying hens reared under high stocking density. Hence, dietary supplementation with GO and VE could have a synergistic positive effect on hen’s performance specifically by minimizing oxidative stress. As such, these compounds can improve nutrient digestibility and consequently enhance productivity during stressful periods. It is important to conduct further research in order to confirm and better understand the specific mechanisms whereby these nutrients have a positive impact on the animal.
The levels of mRNA transcription were confirmed using quantitative real-time RT-PCR. In our results, a significant effect of low stocking density on the expression levels of HMGCR, HSP70, IL-12β, and IFN-γ were not observed among the 4 dietary groups. However, the expression levels of HMGCR and IFN - γ in the blood were significantly impacted by the high stocking density condition, whereas there was no significant difference for HSP70 and IL-12β. The study of Sohn et al. [31] reported greater abundance of HMGCR, but not HSP70, the blood of chickens exposed to stress. A similar finding was reported by Incharoen et al. [11] where HMGCR abundance was lower in laying hens reared at high stocking density fed with dietary germinated paddy rice containing several bioactive compounds (vitamins, GO, and γ-amino butyric acid) [52]. The lower HMGCR abundance in the hens housed under high stocking density that received GO alone or combined with VE could be taken as indication of a reduction in stressful conditions as reported by Sohn et al. [31]. This idea is further supported by data from Zavoshy et al. [53] where feeding vitamin E isomers (tocopherol and tocotrienols) and GO (also contained in rice bran oil) led to lower total cholesterol and low-density lipoprotein levels by inhibiting HMG-CoA reductase, i.e., the rate-limiting enzyme in de novo cholesterol synthesis. Thus, based on our findings, providing dietary GO and VE may have a mitigating effect on the stress status caused by high stocking density conditions.
IFN-γ is a vital cytokine synthesized primarily by type 1 T helper cells and plays a crucial role in the activation of macrophages [54,55]. In avian species, IFN-γ represents a natural component of the immune system [56] and its abundance has been detected in laying hen [11], duck [57] and goose [58]. Thus, the lower abundance of IFN-γ [59] in broiler chickens exposed to heat stress or in birds raised in environmental conditions with higher endotoxin levels underscore the usefulness of this cytokine as a marker of stressful conditions in avian species [60]. Despite the lack of differences in the abundance of IFN-γ due to feeding diets under low stocking density conditions, the fact that dietary GO in hens raised in an environment with high stocking density led to greater IFN-γ mRNA abundance suggests that nutrition may play a role in the function of this cytokine. Lee et al. [61] noted that high levels of IFN- γ have been associated with protective immune responses to parasitic infections. In fact, Gao et al. [57] suggested that IFN-γ has the potential to inhibit viral activity in ducks. Thus, the greater mRNA abundance of IFN-γ in birds fed dietary GO suggests that this compound might aid in mitigating the detrimental impacts of the environment with high stocking density by enhancing the immune response.
CONCLUSION
Hens housed at a 420 cm2/hen high stocking density had lower productivity and eggshell hardness compared with hens housed in an environment with low stocking density (840 cm2/hen). However, dietary GO and VE at a level of 200 ppm, either individually or in combination can improve egg productivity and eggshell hardness as well as regulate mRNA abundance of immune- and stress-related genes. Thus, we conclude that these dietary antioxidants should be part of a nutritional strategy to mitigate the negative impacts on laying hens reared under high stocking density conditions.