INTRODUCTION
Pigs are immunologically and physiologically immature in their weaning phase, associated with decreased growth performance, and an increased risk of infection and disease, particularly diarrhea [1]. Antibiotic growth promoters (AGPs) are added to diets to alleviate weaning stress, increase feed intake and carcass weight, and regulate gut microbial composition [2]. However, growing concerns about the potential negative effects of adding AGPs to pig diets have led to increased interest in raising pigs without the use of AGPs. The removal of AGPs from the diets of freshly weaned pigs may result in health problems and slow their growth [3]. Consequently, management and dietary approaches may need to be modified to avoid the adverse effects of removing AGPs from pig diets.
One of the most promising substitutes for antibiotics is the administration of high doses of zinc (Zn), especially as zinc oxide (ZnO), which is widely commercialized in many nations [4]. ZnO supplementation enhanced piglets’ growth efficiency, digestion, and feed intake, while reducing post-weaning diarrhea and mortality [4–6]. Since it has an antibacterial function, high dosages of ZnO have been found to lower bacterial populations in the intestine and encourage the growth of enterotoxigenic enterobacteria, which alleviates diarrhea [7]. Moreover, ZnO therapeutic dosages (2,500–3,500 mg/kg) decreased diarrhea and improved growth responses in pigs [8], and decreased the production of noxious gases [9]. However, excessive ZnO supplementation (2,500–3,000 ppm) may disrupt gut microbiota balance, leading to increased proliferation of Escherichia coli, which can contribute to diarrhea rather than mitigate it [10]. Moreover, Hahn and Baker [11] reported that long-term consumption of pharmaceutical dosages of dietary ZnO may have several adverse effects, including antibiotic resistance, reduced nutritional absorption, poor growth efficiency, and pollution of the environment. Moreover, prolonged ZnO supplementation at high levels can also induce toxicity and adverse physiological reactions [12]. Therefore, providing ZnO at a substantially lower concentration could be a more suitable approach to achieving the desired benefits while potentially minimizing any associated risks or adverse effects. Hollis et al. [13] reported that including ZnO in the diet at lower levels of 250 or 500 mg/kg did not effectively promote the growth of weaned pigs. Moreover, Biswas et al. [14] suggested that a lower dose of Zn-aspartic acid chelate (750 ppm) can be used as a growth stimulant and to mitigate environmental pollution in weaned piglets, providing a potential replacement for medicinal ZnO. Consequently, optimizing dietary formulations with lower ZnO levels alongside other functional additives, such as probiotics, could provide a viable strategy for maintaining pig health and performance while addressing environmental concerns. Therefore, developing an optimized dietary formula as an alternative to excessive standard ZnO supplementation is essential for improving swine production while ensuring sustainability and minimizing potential risks.
Probiotics have been widely recognized for their ability to enhance growth efficiency, improve nutrient digestibility, and reduce diarrhea incidence in weaned pigs [15]. When incorporated into pig diets, probiotics can exert beneficial effects on gut microbiota by promoting the proliferation of beneficial bacteria, enhancing intestinal barrier function, and modulating immune responses [15,16]. In addition to their role in improving disease resistance, probiotics have been shown to decrease fecal toxic gas emissions, thereby contributing to better air quality and reduced environmental pollution [16]. Notably, dietary supplementation with 1,200 ppm ZnO in combination with probiotics has been reported to yield comparable outcomes to pharmaceutical ZnO levels (3,000 ppm) in terms of growth performance, nutrient utilization, gut microbiota balance, noxious gas emissions, and fecal characteristics in piglets [17]. Furthermore, Shi et al. [9] found that low-dose ZnO combined with probiotic substances improved the growth rate in weaned pigs without impairing immunological parameters, nutritional absorption, or the incidence of gastroenteritis. There has been substantial research on the gut microbiome to reduce the discharge of malodorous substances and improve livestock production and animal welfare [18]. Alterations in the gut microbiota and its byproducts can influence pig health, potentially affecting the animals’ growth performance positively or negatively [19]. Changes in the gut microbiota were noted by boosting beneficial bacteria and diminishing detrimental ones following the addition of probiotics to the pig’s diet [20]. However, despite increasing research interest, there remains a lack of comprehensive data on the combined effects of ZnO and probiotics on the gut microbiome in pigs.
We hypothesized that a reduced level of ZnO combined with probiotics might synergistically improve growth rates, regulate the gut microbiome, boost immune system indicators, and diminish the occurrence of diarrhea and the emission of detrimental gases in pigs. The purpose of this experiment was to examine the impacts of supplementing feed with ZnO and probiotics on productivity, fecal index, meat quality characteristics, odorous substances, and microbiome analysis in weaning-finishing pigs.
MATERIALS AND METHODS
The probiotics complex employed in this study was obtained from a commercial organization, such as SynerZymeF10 from SynerBig. This substance is a combination of spray-dried spores of Bacillus coagulans, Bacillus licheniformis, Bacillus subtilis, and Clostridium butyricum, and it is assured to comprise at least 1 × 1012 CFU/kg B. coagulans, 5 × 1011 CFU/kg B. licheniformis, 1 × 1012 CFU/kg B. subtilis, and 1 × 1011 CFU/kg C. butyricum.
Two hundred weaned piglets from a tertiary hybrid ([Landrace × Yorkshire] × Duroc) were introduced; their initial weight averaged 6.65 ± 0.66 kg, and the feeding trial lasted for 154 days (22 weeks). The test included four treatments: 1) TRT1, basal diet + ZnO 75 ppm; 2) TRT2, basal diet + ZnO 75 ppm + probiotic 0.3% → basal diet + ZnO 75 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%; 3) TRT3, basal diet + ZnO 2,500 ppm → basal diet + ZnO 2,500 ppm → basal diet + ZnO 75 ppm; 4) TRT4, basal diet + ZnO 2,500 ppm + probiotic 0.3% → basal diet + ZnO 2,500 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%, TRT1 was consistent for all and variations in ZnO and probiotic doses in TRT2, TRT3, and TRT4 were applied phase-wise (weaning → growing → finishing). There were 10 repetitions per treatment with each repetition consisted of 5 heads (3 gilts and 2 barrows per repetition), randomly assigned. The trial was distributed into 3 stages 1st as weaning stage (0–6 weeks), 2nd as growing stage (6–12 weeks), and 3rd as finishing stage (12–22 weeks).
The feed was formulated using a corn-soybean meal base and adhered to the nutritional specifications outlined in the NRC [21] (Tables 1, 2, and 3). The test feed, prepared with a feed mixer DDK-801 (Daedong Tech) at the farm, was allowed for free consumption through a one-sided stainless steel self-feeder. Water was provided through an automatic nipple drinker, adjusted for free access. All pigs were kept in climate-controlled pens constructed from stainless steel and featuring slatted plastic flooring. The ambient conditions in the room were maintained at 25°C for temperature and 60% for humidity.
Provided per kg diet: Fe, 100 mg as ferrous sulfate; Cu, 17 mg as copper sulfate; Mn, 17 mg as manganese oxide; I, 0.5 mg as potassium iodide; and Se, 0.3 mg as sodium selenite.
Provided per kg diet: Fe, 100 mg as ferrous sulfate; Cu, 17 mg as copper sulfate; Mn, 17 mg as manganese oxide; I, 0.5 mg as potassium iodide; and Se, 0.3 mg as sodium selenite.
Provided per kg diet: Fe, 100 mg as ferrous sulfate; Cu, 17 mg as copper sulfate; Mn, 17 mg as manganese oxide; I, 0.5 mg as potassium iodide; and Se, 0.3 mg as sodium selenite.
Individual measurements for weight gain per day, daily feed intake, and feed efficiency were taken at the start, weeks 6, 12, and 18, and at the end of the test (week 22). Feed intake was determined by subtracting the remaining amount from feed intake at the time of weight measurement, and feed efficiency was calculated by dividing daily weight gain by feed intake.
It was measured at the beginning of the test, weeks 6, 12, 18, and at the end of the test (week 22), and was calculated by quantifying the following index and averaging it every week (Score: 1 = hard, dry pellets in a small, hard mass; 2 = hard, formed stool that remains firm and soft; 3 = soft, formed, and moist stool that retains its shape; 4 = soft, unformed stool that assumes the shape of the container; 5 = watery, liquid stool that can be poured). Two trained evaluators conducted the assessment in a treatment-blind manner, recording scores per pen based on observed fecal characteristics in specific pigs within the pen [14].
Fresh feces were gathered from two randomly chosen pigs (one gilt and one barrow) in each pen to analyze noxious gases at weeks 6, 12, 18, and 22 for each test, 300 grams of feces were transferred into a 2.6 mL sealed plastic container with a small aperture and left to ferment for 24 hours at 25°C. A hundred-milliliter specimen was detached from the headspace (about 2.0 cm) to permit for airborne movement. After resealing, the container underwent a 30-second manual shaking to inspect the formation of a crust on the surface. NH3, H2S, methyl mercaptan, acetic acid, and CO2 levels were measured utilizing a multi-gas meter (MultiRAE Lite model PGM-6208, RAE Systems). Several measurement tubes (No. 3L, No. 4LT, and No. 70L; Gastec) were utilized to determine total mercaptans. The procedure is implemented in agreement with the procedure outlined by Biswas et al. [22].
From each pen, two pigs were randomly chosen (total of 20 pigs per treatment), weighed, slaughtered through trained personnel, and then examined for meat quality. The pork used for the analysis was stored in a refrigerator at 4°C for 24 hours after slaughter. Subsequently, the M. longissimus dorsi (semiconductor sirloin) was separated and used for further examination. Following the method outlined by Balasubramanian et al. [23], we assessed sensory attributes (color, marbling, and firmness), as well as lightness (L*), redness (a*), yellowness (b*), pH, longissimus muscle area (LMA), cooking loss, and drip loss. To analyze water-holding capacity (WHC), a 0.3 g sample of pork meat was placed on a 125 mm diameter filter paper and pressed for 3 minutes at 26°C. The moisture exposure of the compressed areas was measured using a digital area-line sensor (MT-10S, M.T. Precision). The water ratio in the meat area was subsequently calculated, where a smaller ratio signifies a higher WHC.
In addition to other samples, fecal microbiome analysis was performed by collecting fecal samples via anal massage at 10 and 40 days after weaning and before shipment. The collected samples were stored in a microbiome-specific kit for further analysis. Genomic DNA was extracted using the QIAamp Power Fecal DNA Kit (Qiagen), following the manufacturers protocol. DNA concentration and purity were assessed using a UV spectrophotometer (Mecasys). The V3–V4 hypervariable region of the 16S rRNA gene was amplified and sequenced on an Illumina MiSeq platform (Illumina). Raw sequencing data were processed using the Quantitative Insights into Microbial Ecology (QIIME2, version 2022.8) pipeline [24]. Data pre-processing and quality control were performed using the ‘cutadapt’ and DADA2 plugins [25]. Phylogenetic diversity analysis was conducted within QIIME2. For taxonomic classification, a feature classifier was trained using the SILVA 138_99 database within the ‘q2-feature-classifier’ module of QIIME2 [26]. Principal coordinate analysis (PCoA) was conducted using the ‘q2-diversity’ plugin with the Bray-Curtis distance matrix. Differential taxonomic markers were identified using the “run_lefse” package in the R software, applying the Linear Discriminant Analysis Effect Size (LEfSe) method [27]. In addition, alpha diversity indices (Chao1, Shannon index, Simpson index, Pielou’s evenness, and Observed OTUs), PCoA plots, and relative abundance bar graphs were generated using the ‘ggplot2’ package in R (version 13.8, SILVA 138v) [28].
The variables were statistically analyzed using one-way ANOVA within a randomly selected complete block design, with feeding strategies as the classification variable. Duncan’s multiple comparison tests were performed to determine if there were substantial alterations between the means. The pen served as the experimental unit, and the SEM was used to represent data variability. Significant differences were indicated by p < 0.05, while p < 0.10 was considered as a trend. Permutational multivariate analysis of variance (PERMANOVA) was utilized to evaluate the significance of the PCoA plot.
RESULTS
According to Table 4, the TRT2 and TRT4 groups presented a higher (p < 0.05) body weight (BW) than the TRT1 and TRT3 at 18 and 22 weeks of the test. Pigs in a TRT2 and TRT4 treatment group had superior average daily gain (ADG) (p < 0.05) at weeks 18, 22, and overall than the TRT1 and TRT3 treatmant groups. Furthermore, ADG tended to enhance (p < 0.10) at week 12 for TRT4 group compared to TRT1. Additionally, average daily feed intake (ADFI) was greater (p < 0.05) in the TRT2 group in the overall period compared to the TRT1 and TRT3 groups. However, the dietary treatment showed no noteworthy impact (p > 0.05) on feed conversion ratio (FCR) throughout the experiment.
ADG, average daily gain; ADFI, average daily feed intake; FCR, feed conversion ratio. TRT1, basal diet + ZnO 75 ppm; TRT2, basal diet + ZnO 75 ppm + probiotic 0.3% → basal diet + ZnO 75 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%; TRT3, basal diet + ZnO 2,500 ppm → basal diet + ZnO 2,500 ppm → basal diet + ZnO 75 ppm; TRT4, basal diet + ZnO 2,500 ppm + probiotic 0.3% → basal diet + ZnO 2,500 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%.
Table 5 describes the outcome of ZnO and probiotic addition in feed on fecal indices of wean-finishing pigs. During the entire test period, fecal indices did not differ significantly among the treatment groups (p > 0.05).
Fecal score = 1 hard, dry pellet; 2 firm, formed stool; 3 soft, moist stool that retains shape; 4 soft, unformed stool that assumes shape of container; 5 watery liquid that can be poured.
TRT1, basal diet + ZnO 75 ppm; TRT2, basal diet + ZnO 75 ppm + probiotic 0.3% → basal diet + ZnO 75 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%; TRT3, basal diet + ZnO 2,500 ppm → basal diet + ZnO 2,500 ppm → basal diet + ZnO 75 ppm; TRT4, basal diet + ZnO 2,500 ppm + probiotic 0.3% → basal diet + ZnO 2,500 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%.
Table 6 shows the effects of ZnO and probiotic inclusion in the feed on the odorous substances of weaning-finishing pigs. At the 18th and 22nd weeks of the test, the TRT2 and TRT4 groups exhibited considerably inferior (p < 0.05) NH3 and H2S levels than the TRT1. Nevertheless, there was no noteworthy impact from the treatment observed (p > 0.05) on acetic acid, R-SH, and CO2 gas emissions throughout the entire testing period.
NH1, ammonia; H1S, hydrogen sulfide; CO1, carbon dioxide. TRT1, basal diet + ZnO 75 ppm; TRT2, basal diet + ZnO 75 ppm + probiotic 0.3% → basal diet + ZnO 75 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%; TRT3, basal diet + ZnO 2,500 ppm → basal diet + ZnO 2,500 ppm → basal diet + ZnO 75 ppm; TRT4, basal diet + ZnO 2,500 ppm + probiotic 0.3% → basal diet + ZnO 2,500 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%.
Table 7 illustrates the effects of incorporating ZnO and probiotics into the diet on the meat quality traits of weaned-finishing pigs. There was no significant change among treatments regarding pH, water holding capacity, cooking loss, longissimus muscle area, drip loss, meat color, and sensory evaluation of meat quality characteristics (p > 0.05).
TRT1, basal diet + ZnO 75 ppm; TRT2, basal diet + ZnO 75 ppm + probiotic 0.3% → basal diet + ZnO 75 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%; TRT3, basal diet + ZnO 2,500 ppm → basal diet + ZnO 2,500 ppm → basal diet + ZnO 75 ppm; TRT4, basal diet + ZnO 2,500 ppm + probiotic 0.3% → basal diet + ZnO 2,500 ppm + probiotic 0.1% → basal diet + ZnO 75 ppm + probiotic 0.1%.
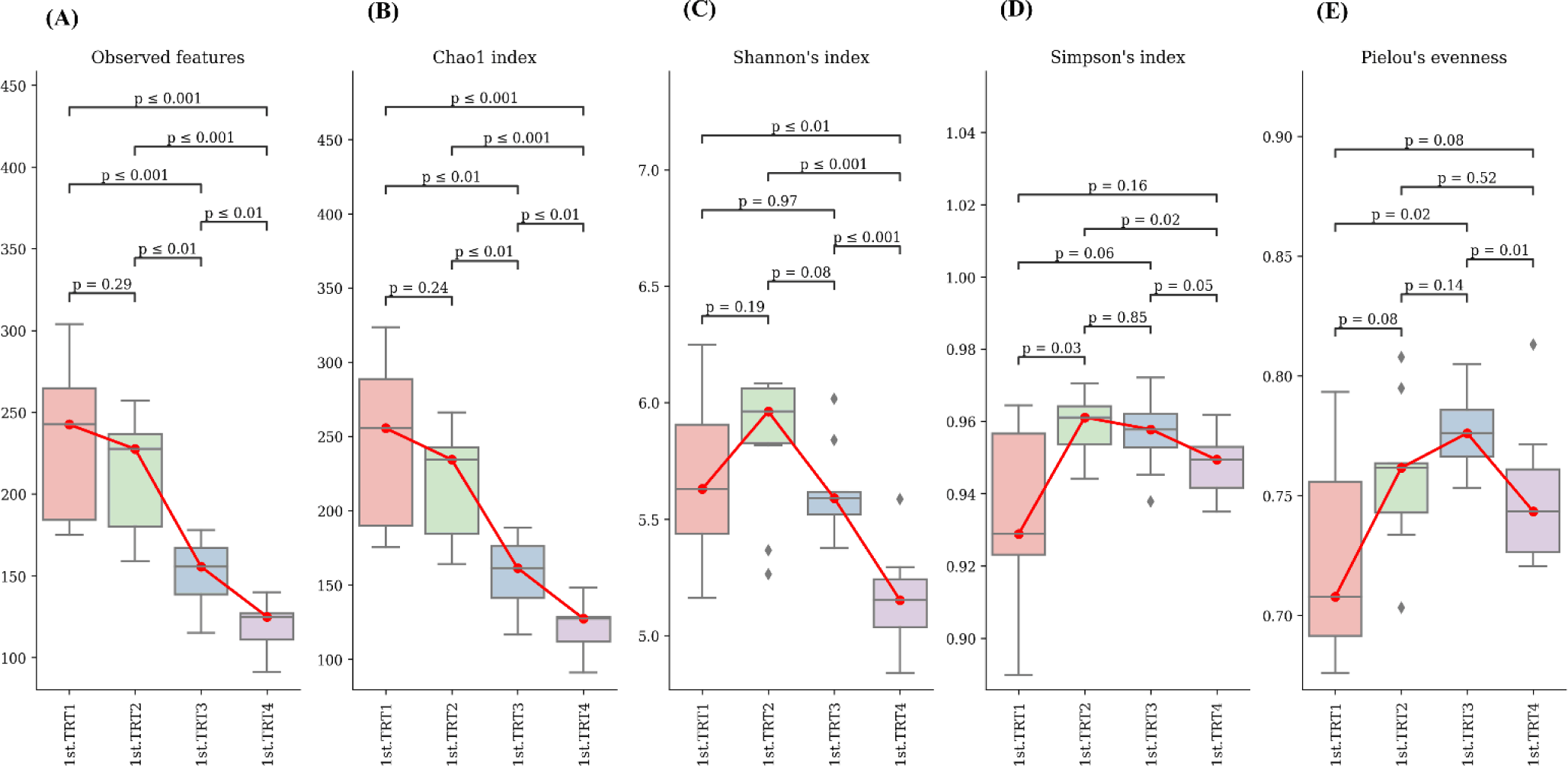
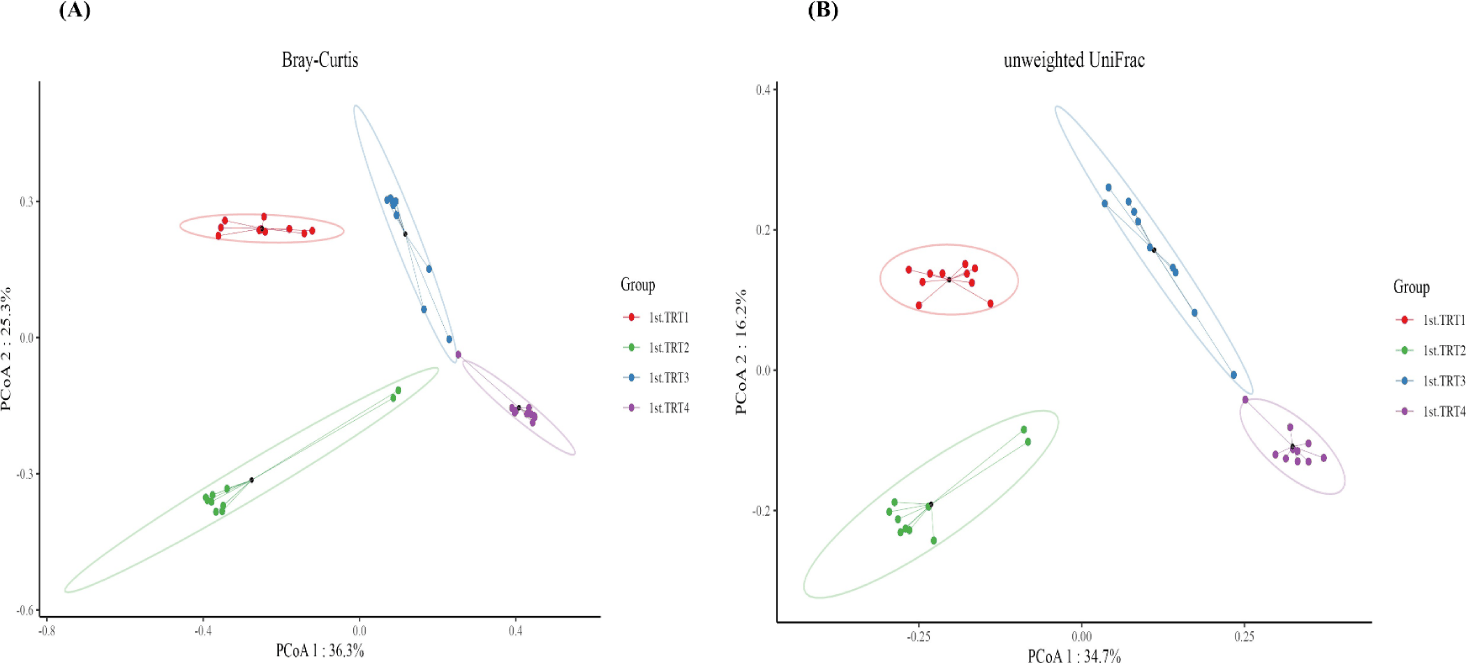
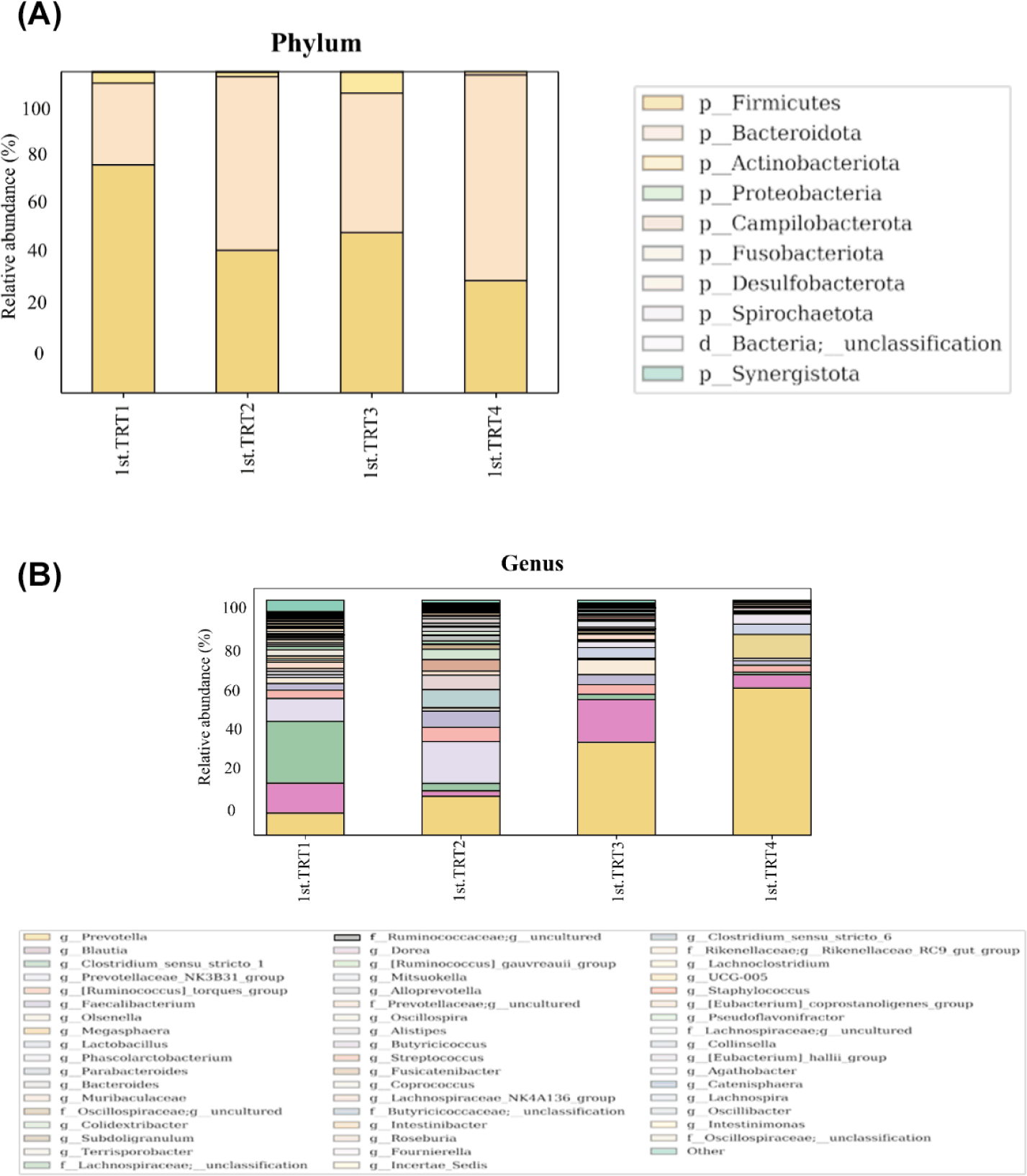
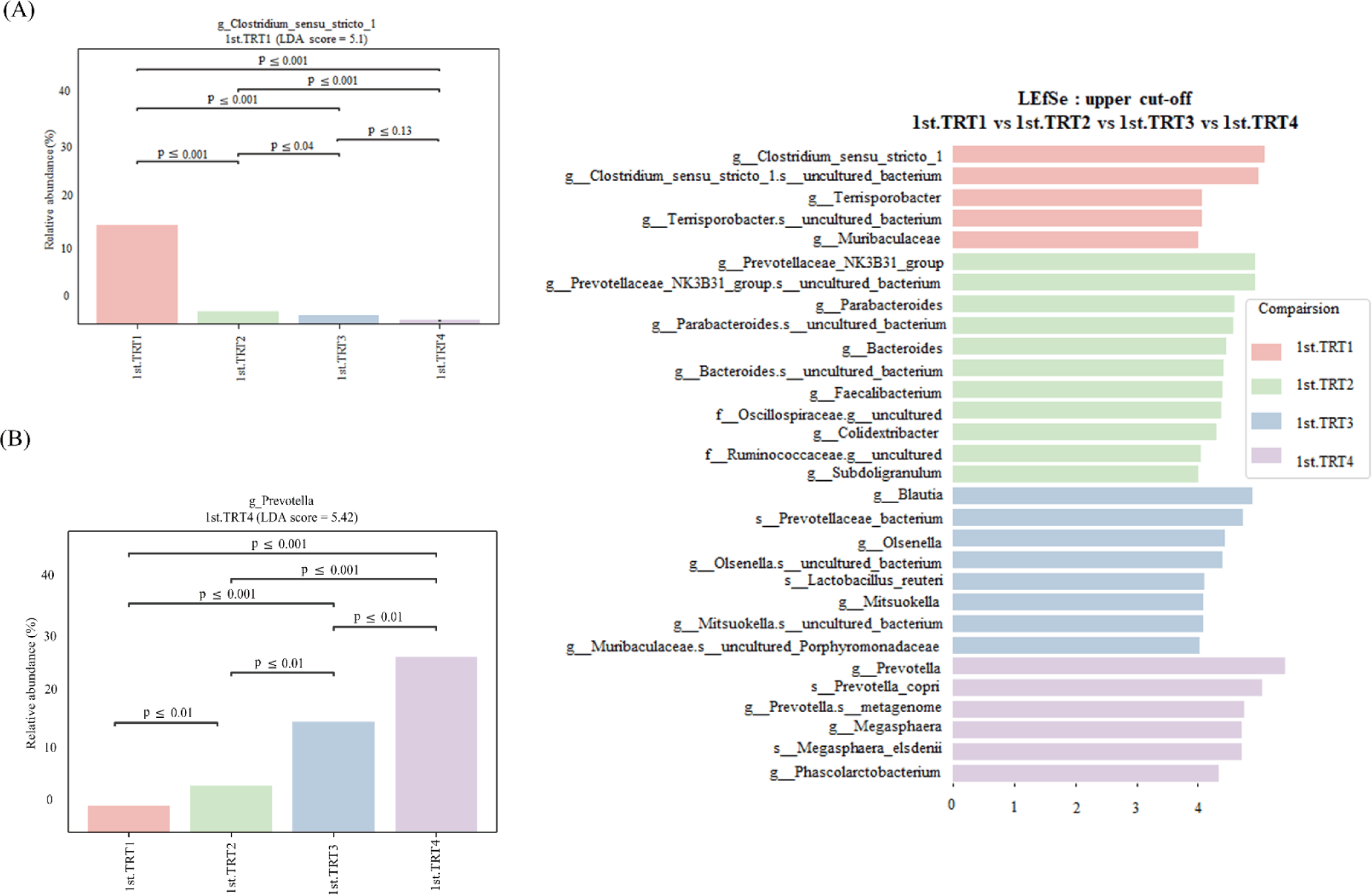
Weaning stage: The alpha diversity indices (Fig. 1) of Chao1, the number of observed features, and Shannon’s index were significantly lowers (p < 0.05) in the TRT4 group compared to other treatment groups. Moreover, the Chao1 index and observed features of the TRT3 group were lower (p < 0.05) than the TRT1 and TRT2. The Simpson’s index was notably greater (p < 0.05) in the TRT2 group in comparison to TRT1, while lower (p < 0.05) in the TRT4 group than the TRT2. In comparison with the TRT1 group, Pielou’s evenness was higher (p < 0.05) in the TRT3, while lower (p < 0.05) in the TRT4 group compared to the TRT3. The gut microbiota of the probiotic and ZnO-treated pigs showed substantially different clusters from that of the TRT1 pigs, according to the PCoA plot generated with the Bary-Curtis and unweighted UniFrac dissimilarity index (Fig. 2). Within the groups, we recognized ten prevailing species at the phylum level (Fig. 3A; with an abundance cut-off set at 0.1%). Firmicutes and Bacteroidota, accounting for 97% of all sequence reads, appeared as the dominant phyla in all the groups. The inclusion of ZnO and probiotics resulted in a significant reduction in Firmicutes levels (p < 0.05) while concurrently increasing Bacteroidota abundance (p < 0.05). In the genus level, enhanced (p < 0.05) Prevotella, and decreased Blautia, Clostridium sensu stricto 1, and Prevotellaceae NK3B31 group were observed (p < 0.05) in the TRT4 group in comparison to the TRT1 (Fig. 3B). The dietary TRT4 treatment group significantly decreased (p < 0.05) Clostridium sensu stricto 1 bacteria and increased (p < 0.05) Prevotella bacteria compared to others, according to LEfSe analysis (Fig. 4). The TRT2 group had lower Prevotella bacteria than TRT3 (p < 0.05) but higher than TRT1, and TRT3 had higher Prevotella bacteria than TRT1 (p < 0.05). Moreover, Clostridium sensu stricto 1 bacteria were lower (p < 0.05) in the TRT2 group than in the TRT1 group, but higher than in the TRT3 group; they were also lower (p < 0.05) in the TRT3 group than in the TRT1 group.
Growing stage: The alpha diversity metrics (Fig. 5), including the count of observed features and the Chao1 index, showed notably reduced (p < 0.05) values in the TRT4 group than the TRT1. Moreover, Shannon’s index, Simpson’s index, and Pielou’s evenness were significantly higher (p < 0.05) in the TRT2, TRT3, and TRT4 groups than the TRT1. By contrast, the TRT3 and TRT4 groups had lower (p < 0.05) Shannon’s index, Simpson’s index, and Pielou’s evenness than the TRT2 group. Additionally, the TRT4 group showed greater (p < 0.05) Pielou’s evenness when compared with the TRT3 group. The PCoA plot utilizing the Bary-Curtis and unweighted UniFrac dissimilarity index revealed distinct clustering between the gut microbiota of the probiotic and ZnO-treated pigs compared to the TRT1 pigs (Fig. 6). Supplementing with ZnO and probiotics led to a notable rise (p < 0.05) in Firmicutes and Bacteroidota abundance at the phylum level (p < 0.05) alongside a concurrent decrease (p < 0.05) in Proteobacteria abundance (p < 0.05) (Fig. 7A). At the genus level, Prevotella and Lactobacillus were increased (p < 0.05); in contrast, Succinivibrio and Clostridium sensu stricto 1 were reduced in all treatment groups compared to the TRT1 group (Fig. 7B). A LEfSe analysis was conducted to identify distinctive taxonomic markers within the pig’s gut (Fig. 8). The dietary treatments notably elevated Prevotella bacteria while reducing Clostridium sensu stricto 1 bacteria compared to the TRT1 group. The TRT2 had greater (p < 0.05) Prevotella bacteria than TRT3 and TRT1, and TRT3 had higher Prevotella (p < 0.05) bacteria than the TRT1 group. Moreover, Clostridium sensu stricto 1 bacteria was inferior (p < 0.05) in the TRT2 group than the TRT3 and TRT1 and also lower (p < 0.05) in the TRT3 than the TRT1.
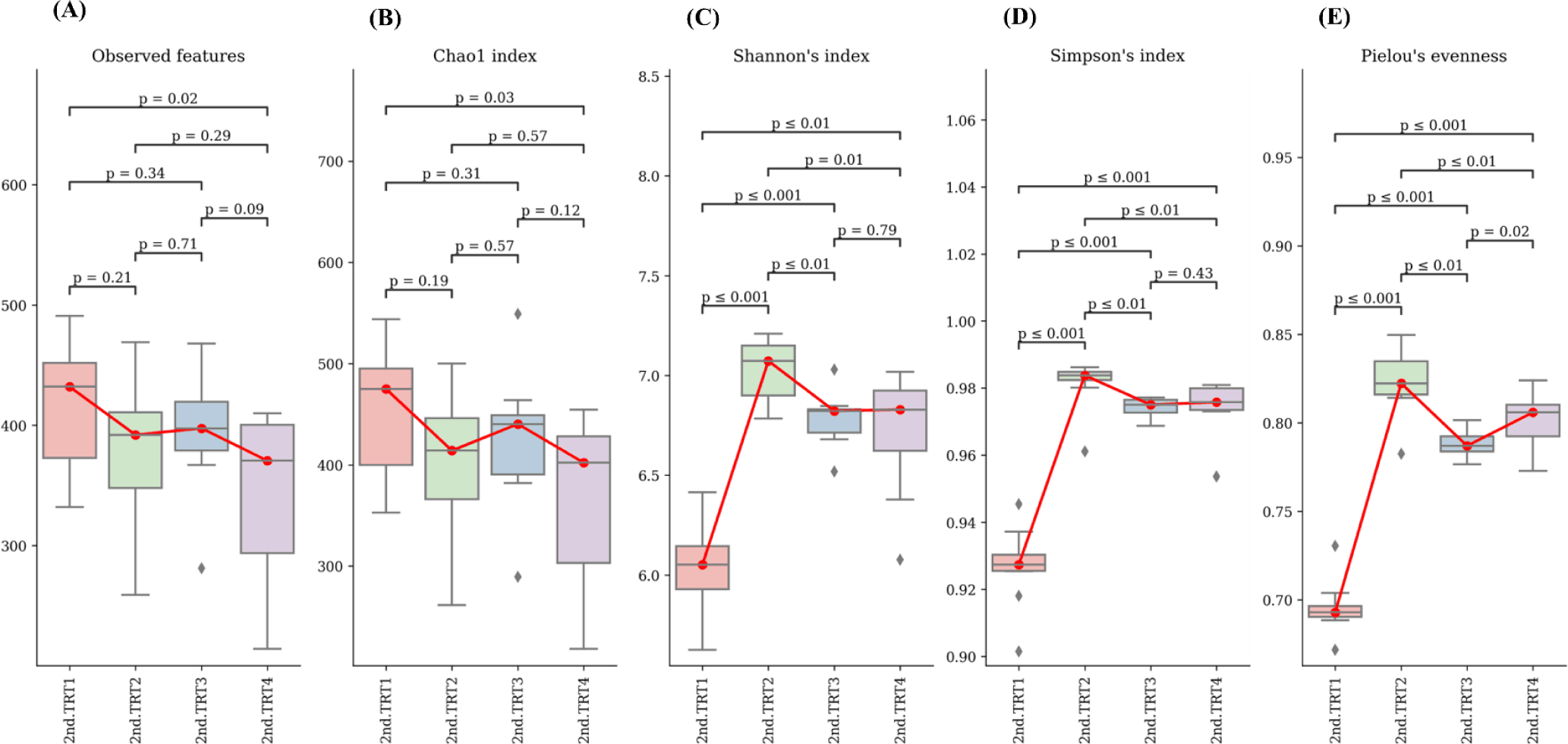
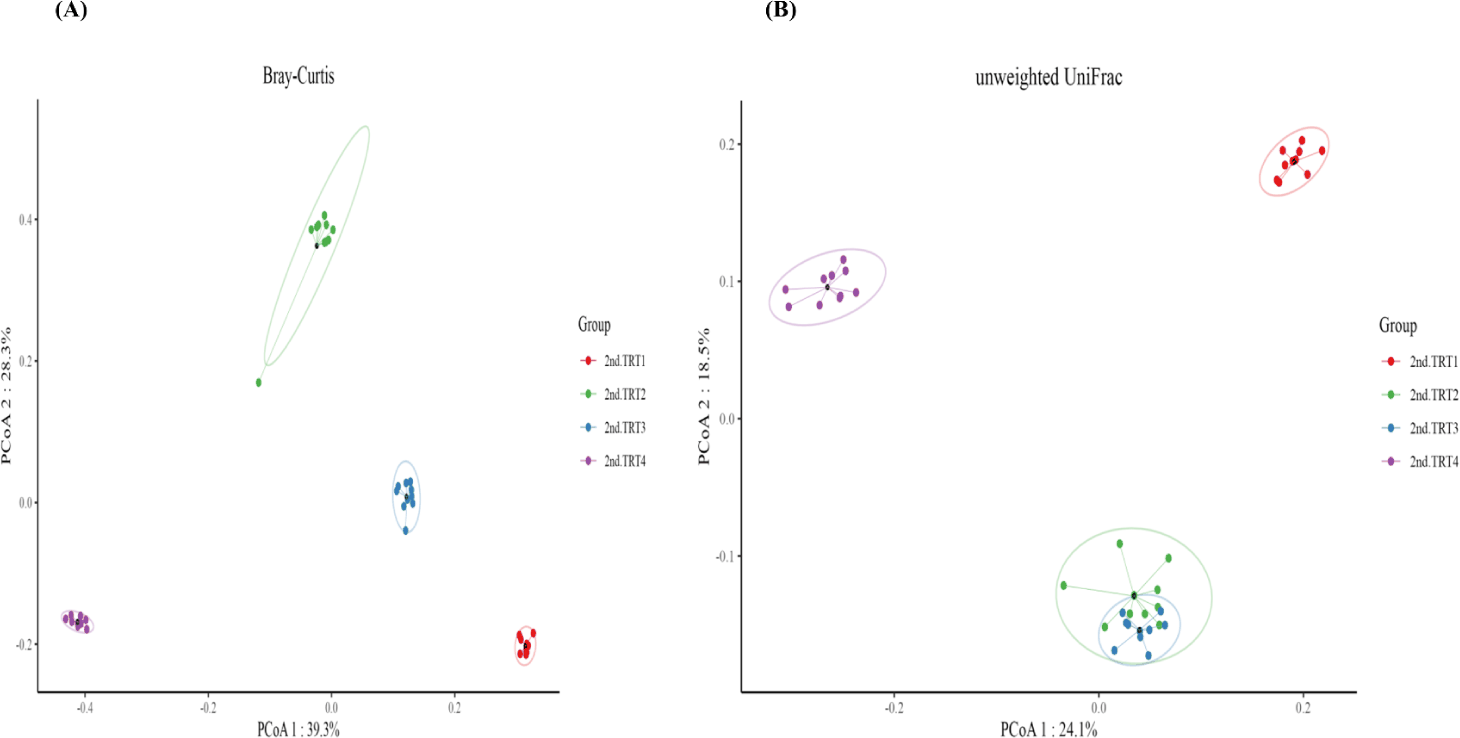
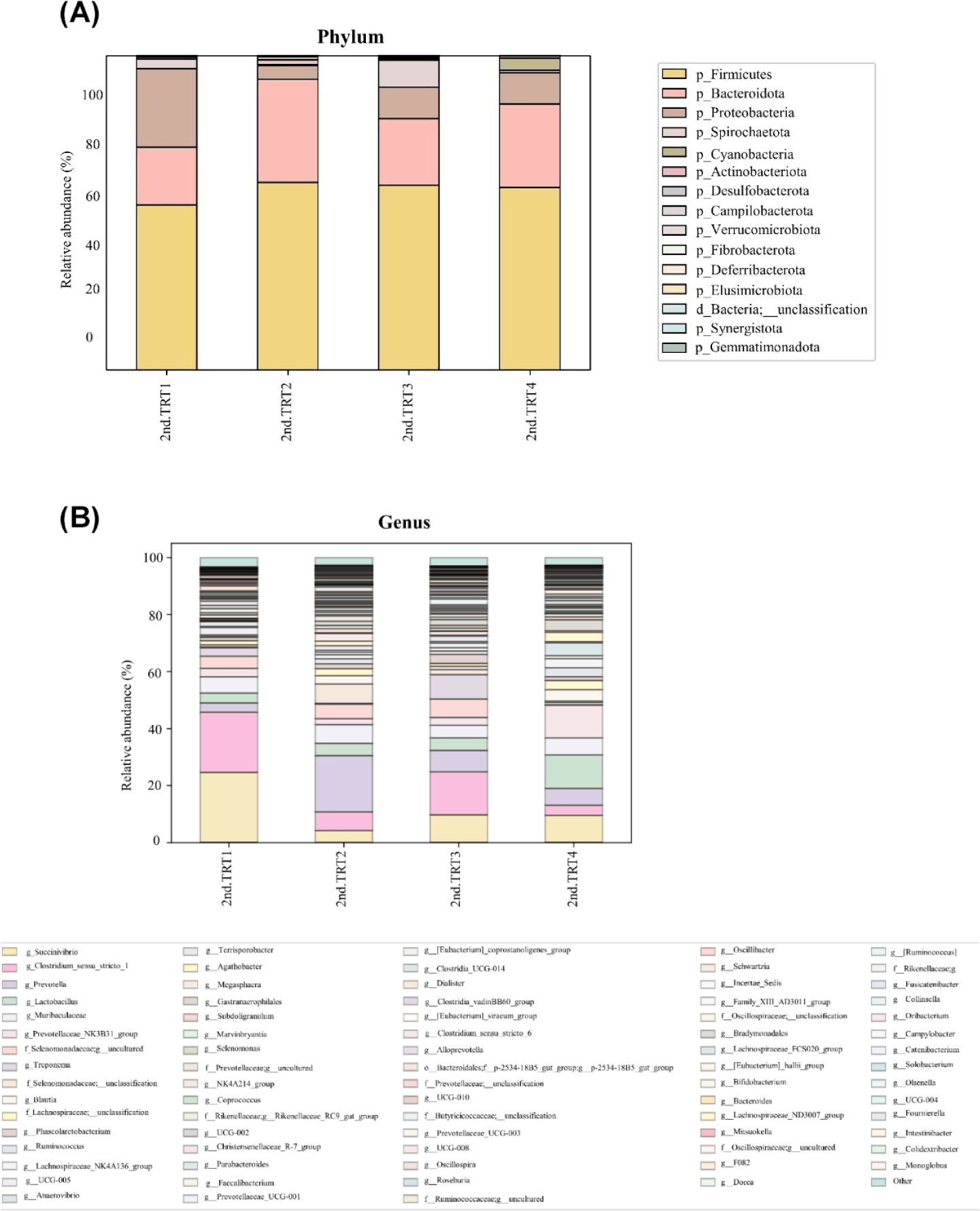
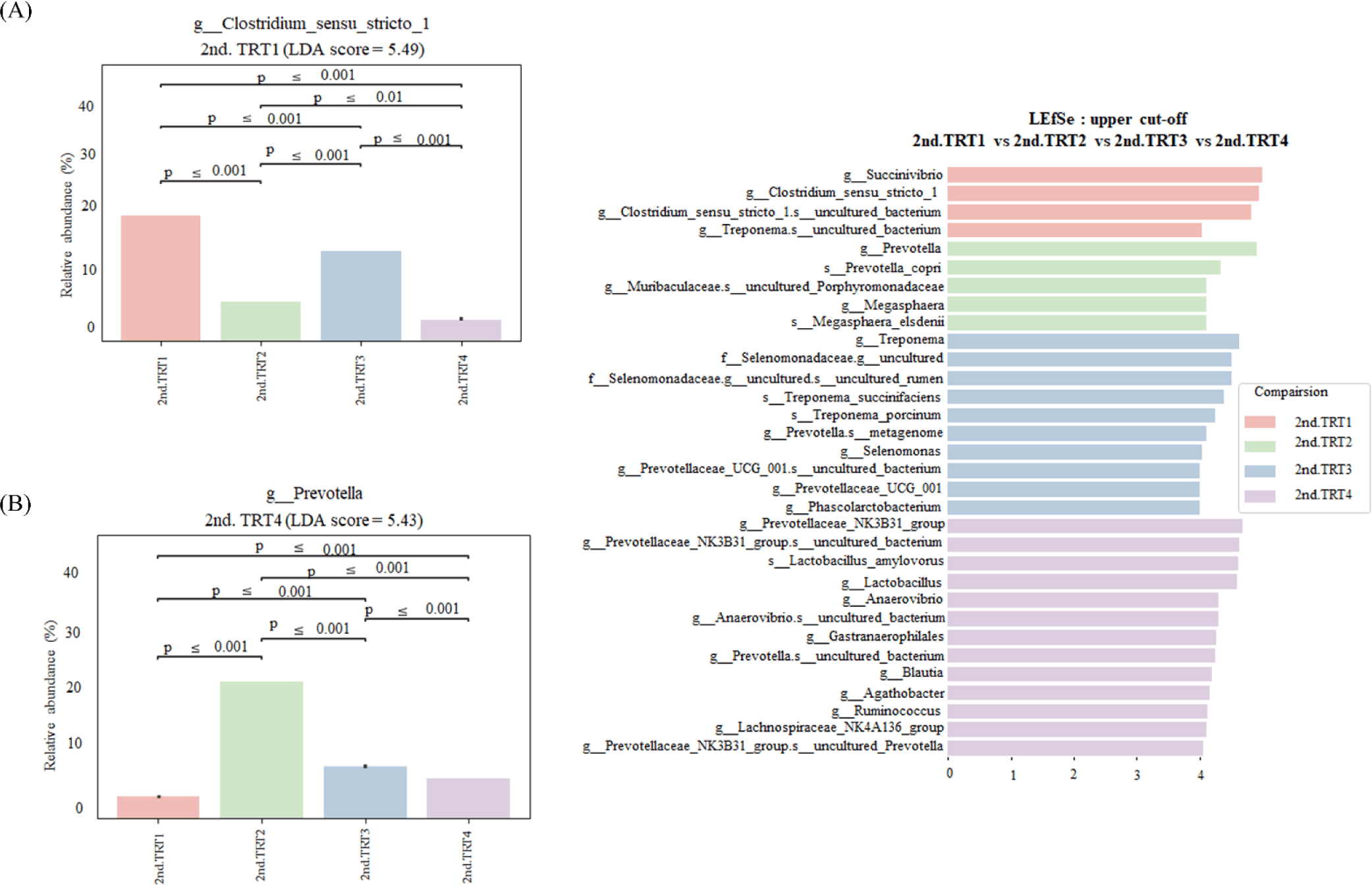
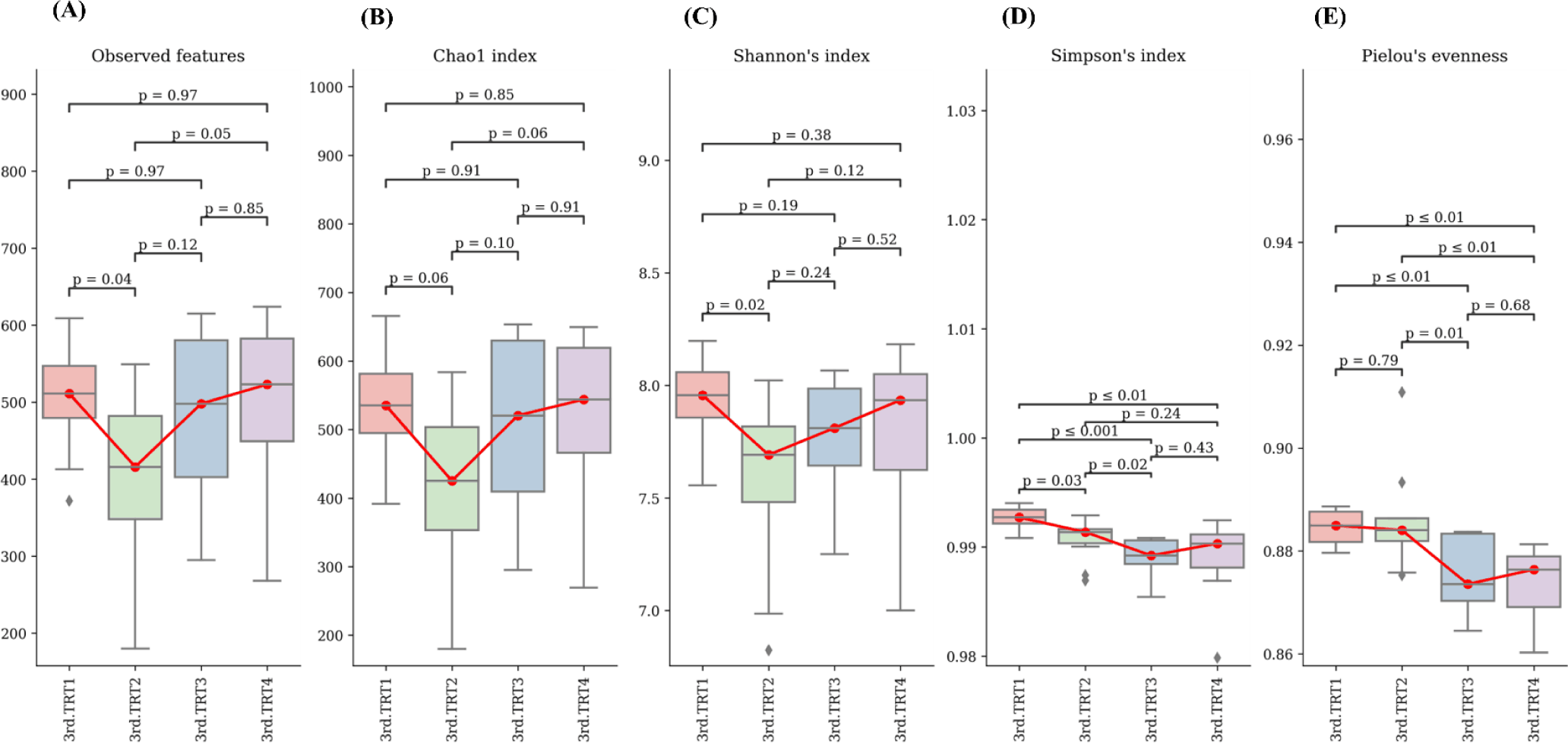
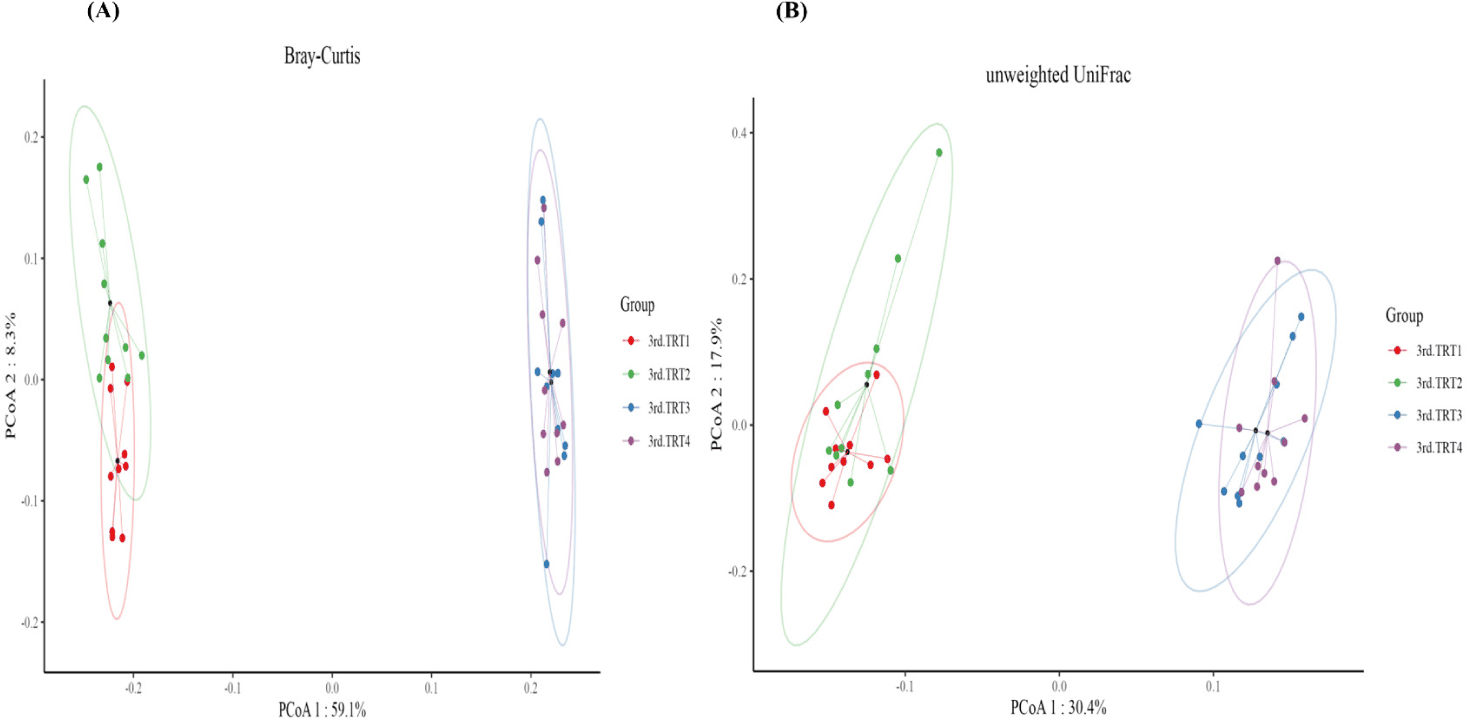
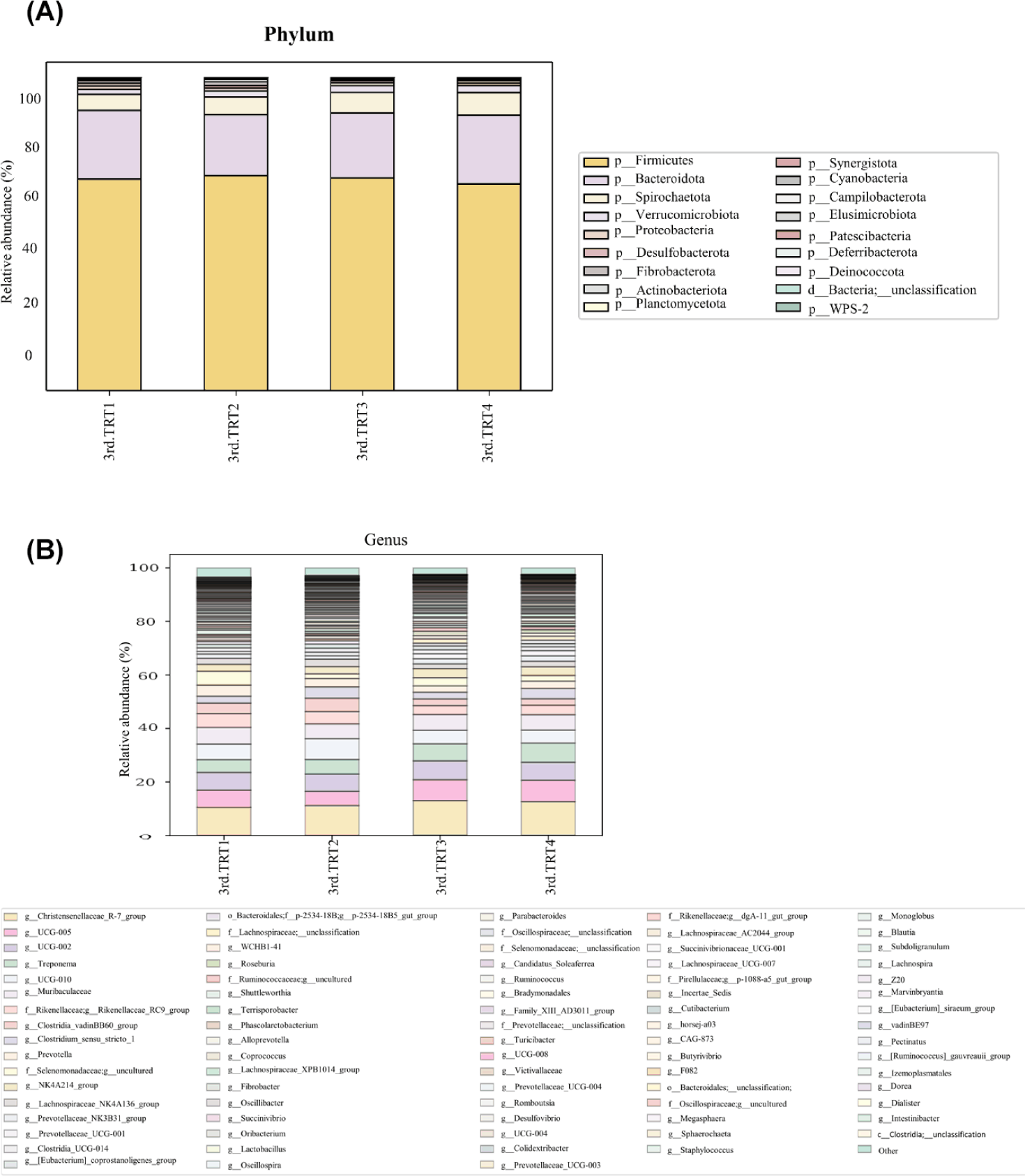
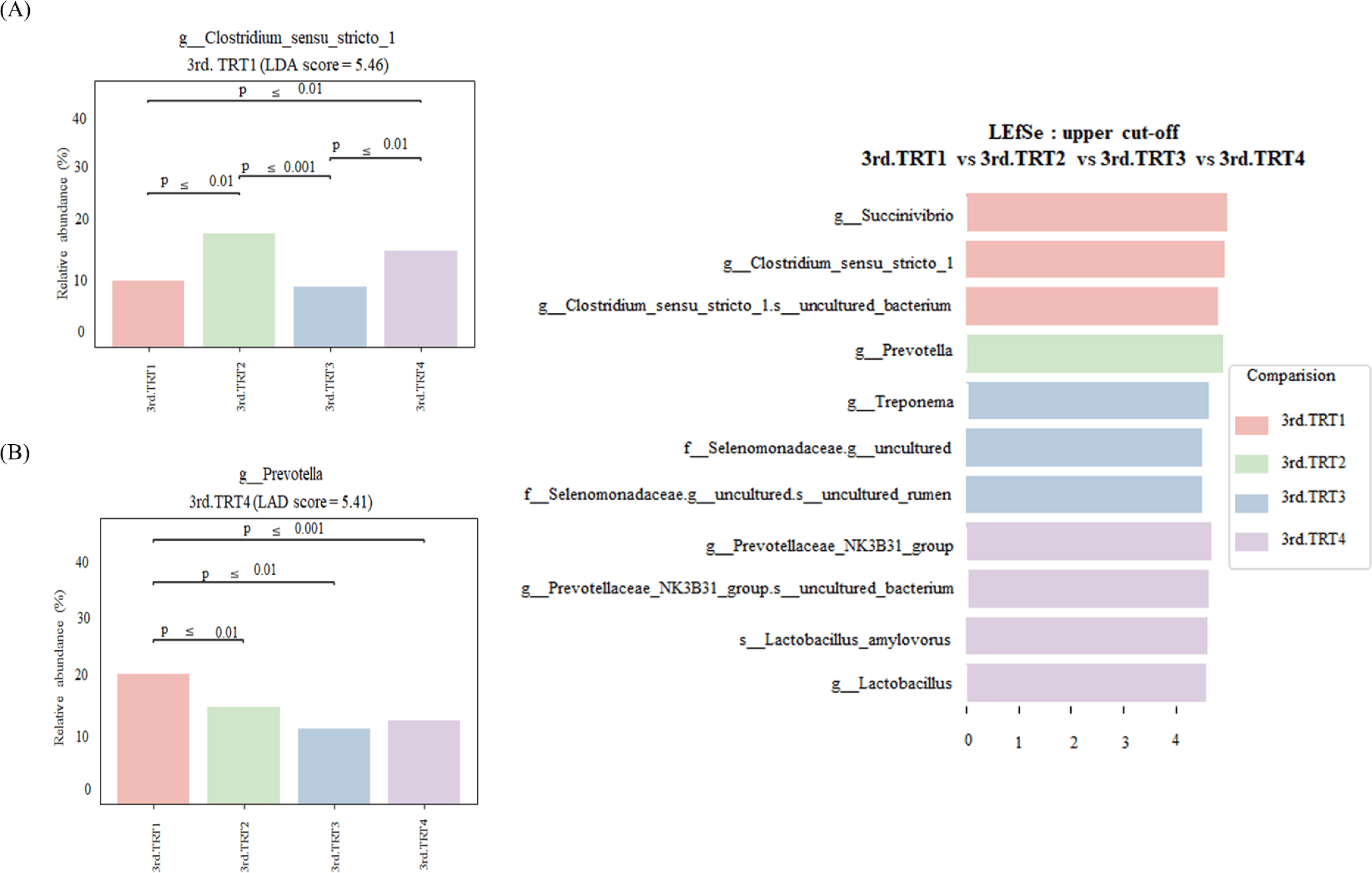
Finishing stage: The alpha diversity measurements (Fig. 9), such as the count of observed features and Shannon’s index exhibited considerably poorer (p < 0.05) values in the TRT2 treatment group when compared to the TRT1. The Simpson’s index was lower (p < 0.05) in all the ZnO-probiotic treated pigs than the TRT1 pigs; additionally lower (p < 0.05) in the TRT3 group than the TRT2 group. The Pielou’s evenness was lower (p < 0.05) in the TRT3 and TRT4 groups than in the TRT1 group. Moreover, the TRT2 group had greater (p < 0.05) Pielou’s evenness than the TRT3 and TRT4 groups. However, Chao1 index revealed no noteworthy difference (p > 0.05). The PCoA plot created based on Bary-Curtis and unweighted UniFrac dissimilarity index demonstrated that the gut microbiota of the ZnO-probiotic included group had considerably distinct clusters from that of the TRT1 pigs (Fig. 10). Firmicutes were the principal taxa at the phylum level, except for Bacteroidota and Spirochaetota in the gut microbiome (Fig. 11A). At genus level (Fig. 11B; relative abundance > 0.1% in all samples), Christensenellaceae R-7 group, UCG-005, UCG-002, and Treponema enhanced; in contrast, UCG-010 and Muribaculaceae reduced in the treatment groups compared to the TRT1 group. The dietary treatments significantly decreased (p < 0.05) Prevotella bacteria in comparison to the TRT1 groups, according to LEfSe analysis (Fig. 12). By contrast, TRT2 and TRT4 had higher (p < 0.05) Clostridium sensu stricto 1 bacteria than the TRT1 group. Moreover, Clostridium sensu stricto 1 bacteria were lower (p < 0.05) in the TRT3 group than TRT2, while higher (p < 0.05) in the TRT4 group than the TRT3 group.
DISCUSSION
Using low-dose ZnO combined with probiotics is valuable as it provides a sustainable approach to improving pig performance and gut microbiota composition while minimizing the adverse effects of high Zn levels on the environment and animal health. The results demonstrated that probiotic supplementation, particularly in combination with lower ZnO levels, significantly enhanced BW, ADG, and FI while reducing NH3 and H2S emissions. Additionally, microbiome analysis revealed a notable increase in beneficial bacteria, alongside a reduction in pathogenic bacteria. These findings align with previous research indicating that probiotics can enhance gut microbial diversity, improve productivity, and contribute to sustainable swine production, while minimizing environmental impact.
The effects of combining probiotics and ZnO in the diets of weaned piglets have been widely investigated, revealing diverse outcomes on productivity and gut health. The TRT2 and TRT4 treatment groups significantly improved BW and ADG at various weeks, with TRT2 also increasing ADFI, but neither treatment affected the FCR throughout the experiment. Li et al. [29] stated that probiotic (Lactobacillus acidophilus) and ZnO administration increased BW and ADG and lowered FCR but showed no substantial interaction between probiotic and ZnO of weaned piglets. Similarly, Shi et al. [9] reported that supplementing diets containing low levels of ZnO (300 mg/kg) with a probiotic complex improved ADG during week 6, while having no effect on BW or the G:F ratio. Compared to the control (CON) diet, the ZnO, benzoic acid, and probiotics mixture substantially enhanced daily FI and ADG, and reduced FCR [30]. Conversely, the administration of ZnO (300 ppm) with 0.1% probiotic lowered ADG compared with CON treatment and decreased the ZnO content resulting in a linear decrease in ADG at phase 3 [17], contrary to our study. However, Biswas et al. [31] stated that dietary incorporation of higher dose ZnO (3,000 ppm) with 0.1% probiotic improved the growth efficiency of weaning pigs compared to other treatment groups, with no synergistic effect between ZnO and probiotic. The discrepancies between these studies could be attributed to alterations in the dosage and mixture of ZnO and probiotics used. For instance, the high dose of ZnO (3,000 ppm) used by Biswas et al. [31] might have provided a stronger antibacterial effect, enhancing gut health and growth performance, whereas lower doses used in other studies may not have been as effective. Additionally, the specific strains of probiotics and their ability to survive and function in the gut environment of pigs can vary, potentially influencing the outcomes. Probiotics are recognized for their ability to modulate the microbiota by promoting the growth of beneficial bacteria, enhancing gut health, and lowering the occurrence of diarrhea [32], which could explain the improved growth capabilities observed in some studies. Meanwhile, the antibacterial properties and improved gut health by ZnO [31] might be the possible cause for improved growth efficiency in our study. The mixed results highlight the complexity of interactions between dietary components and the need for optimizing dosages and combinations to achieve consistent benefits in growth performance and gut health in pigs.
One of the airborne contaminants in current intensive pig production is fecal harmful gas releases, such as NH3 and H2S. Scientists are searching for new feeding strategies to prevent environmental pollution in the swine production industry. Wang et al. [33] observed that administering probiotics like B. subtilis and B. licheniformis significantly reduced slurry NH3 gas emissions in growing pigs, although this treatment had no impact on H2S and R-SH release. Similarly, Lan et al. [34] found that adding a probiotic complex containing B. coagulans, B. licheniformis, B. subtilis, and C. butyricum to a basal diet without additional ZnO decreased NH3 and H2S emissions similar to our research. Chen et al. [35] also revealed that adding Bacillus-based probiotics (0.2%) to the diet decreased NH3 emissions, suggesting that probiotics can positively impact the gut microbial environment, enhance nitrogen digestion, and subsequently decrease excreta noxious releases [36]. Besides, Yan and Kim [37] indicated that the advantageous impact of probiotics on the large intestine’s gut microflora could explain the reduced fecal noxious gas concentration. In contrast, other studies have shown different results. According to Shi et al. [9] and Wang et al. [17], supplementing a diet with both low and high doses of ZnO with a probiotic complex did not affect fecal gas emissions in weaned pigs. Likewise, Biswas et al. [31] found that adding a probiotic complex containing ZnO to the piglets’ diet did not affect their gas emissions. However, our findings differ from these latter studies, as we observed a reduction in NH3 and H2S emissions with dietary probiotic administration. In our study, the decline in gas emissions can be attributed to the beneficial alterations in the gut microbiota. The probiotics used in our dietary approach increased beneficial bacteria such as Firmicutes, Prevotella, and Lactobacillus while reducing harmful bacteria like Clostridium sensu stricto 1 and Bacteroidota. These changes likely enhanced nitrogen digestion and overall gut health, leading to decreased fecal noxious releases. The variation in gas emission results observed in different studies could be due to several factors. One possible reason is that the specific strains and dosages of probiotics used might not have been effective in altering the gut microflora sufficiently to impact gas emissions. Another factor could be the interaction between ZnO and probiotics; ZnO’s antibacterial properties might counteract the advantageous effects of probiotics on gut health and nitrogen metabolism. Additionally, variations in diet composition, environmental conditions, and animal physiology could influence the outcomes, making it difficult to achieve consistent reductions in gas emissions across different studies. These findings emphasize the necessity for future study to optimize the combinations and dosages of probiotics and ZnO to achieve the desired environmental benefits in swine production.
The weaning phase in pigs poses significant concerns due to dietary, environmental, and gut morphological shifts that can result in decreased growth rates, heightened diarrhea incidence, and an inadequate intestinal ecosystem [38]. Biswas et al. [31] found that including dietary probiotics and ZnO in the feed enhanced fecal Lactobacillus and decreased E. coli counts, resulting in no diarrhea incidence in weanling piglets. Similarly, Shi et al. [9] reported no diarrhea among pigs receiving either high or low-level ZnO in combination with probiotics, as all pigs exhibited a fecal score ranging from 3.2 to 3.4. Giang et al. [38] also stated that a Bacillus-based microbiome reduced the frequency of diarrhea and fecal scores in piglets. Additionally, Milani et al. [39] found that piglets given nano-sized ZnO had lower fecal scores than those given a CON diet. However, Wang et al. [17] noted that adding 0.1% probiotics complex with ZnO (3,000 ppm or 1,200 ppm) into the diet had no discernible effect on the fecal score of weanling piglets, similar to our study. The presence of probiotics in the feed has been shown to notably influence intestinal bacterial populations, leading to decreased diarrhea scores [40]. Additionally, ZnO demonstrates the capacity to bolster the gut microbiome, fostering competition among enterobacteria [17]. The discrepancies between studies may be due to differences in the types and dosages of probiotics and ZnO used, as well as variations in diet composition and environmental conditions. In the present study, fecal scores were not significantly altered by ZnO supplementation, suggesting that its effects on gut health may not always be directly reflected in observable fecal consistency. However, ZnO is known to play a crucial role in modulating gut microbiota, intestinal integrity, and immune responses, which are not solely indicated by fecal scoring. The combined effects of probiotics and ZnO on gut health and diarrhea incidence underscore the complexity of optimizing dietary strategies for weanling piglets to improve health outcomes and growth performance.
Finishing pigs receiving ZnO showed enhanced meat quality, as demonstrated by improvements in meat color attributes and reduced cooking loss [41]. Chen et al. [42] stated that dietary Nano-ZnO improved meat quality by reducing drip loss (after 48 hours), cooking loss, and shearing force, particularly in pigs with intrauterine growth retardation. Furthermore, Natalello et al. [43] found that including 45 or 100 mg/kg of Zn-glycine in the diet minimized carcass chill loss, although cold carcass weight did not significantly differ between treatments. Similarly, Balasubramanian et al. [23] detected that nutritional administration of probiotics without ZnO decreased drip loss and cooking loss in growing-finishing pigs, and Meng et al. [44] stated an increase in longissimus muscle area with probiotic supplementation. Conversely, Cernauskiene et al. [45] found that supplementation of the probiotic E. faecium had no appreciable impact on the meat quality of fattening pigs, consistent with the findings of the current investigation. These inconsistent results can be influenced by various experimental parameters, feed compositions, bacterial species used, and pig genotypes [46]. The impact of combining probiotics and ZnO on pig meat quality remains underexplored in the literature, suggesting the need for further research to assess the combined effects of probiotic and ZnO inclusion on meat quality in pigs.
The gut microbiota plays a crucial role in intestinal barrier integrity, the maturation of mucosal functions, the development of the immune system, aiding nutrient absorption, and regulating energy metabolism [47]. In our research, ZnO-probiotic administration resulted in increased richness of alpha diversity metrics (observed features, Shannon, Simpson, and Chao1). These changes can be attributed to the synergistic effects of ZnO and probiotics in creating a more favorable gut environment for diverse microbial populations. Conversely, Cui et al. [48] reported that incorporating B. subtilis into the feed did not significantly impact species richness or diversity in weaned pigs compared to the CON group. However, Ding et al. [49] found that while B. subtilis did not alter the alpha diversity, it did cause notable differences in the microbial composition of the ileum and jejunum contents between the CON and probiotic groups. Similarly, Long et al. [50] observed that ZnO supplementation increased the microbial β-diversity index in both the ileum and colon, although it reduced microbial α-diversity in the ileum and enhanced it in the colon. This recommends that ZnO and probiotics can influence specific gut regions differently, potentially promoting microbial diversity and intestinal health. Mun et al. [51] reported no significant differences in microbial diversity between the CON and probiotic groups based on weighted UniFrac distances and PCoA analysis. Additionally, Deng et al. [52] noted that higher doses of B. subtilis resulted in a clear distinction between groups but did not alter alpha diversity compared to the CON group. In our study, the PCoA plot, utilizing Bray-Curtis and unweighted UniFrac dissimilarity indices, showed distinct clusters between the gut microbiota of ZnO-probiotic treated pigs and the CON pigs. Significant bacterial groups, such as Firmicutes and Bacteroidetes, play vital roles in host health by influencing processes such as fat metabolism and carbohydrate fermentation [53]. Higher levels of Firmicutes are associated with increased energy production, active transport, facilitated diffusion, endocytosis, and passive diffusion, whereas elevated levels of Proteobacteria and Bacteroidetes in fecal samples are linked to reduced nutrient digestibility [54]. Our study’s results align with findings by Zhang et al. [55], showing that probiotic supplementation increased Firmicutes levels and decreased Bacteroidota abundance compared to the CON group. This shift could be due to the enhanced competitive advantage given to beneficial bacteria by the probiotics and ZnO, leading to better nutrient utilization and growth performance. Besides, Guo et al. [56] found that an elevated Firmicutes/Bacteroidetes ratio is typically associated with higher body weight, consistent with our study’s outcomes. Proteobacteria act as a marker for potential gut ailments and are essential in managing growth performance while preserving energy metabolism [57]. Lactobacillus is recognized for its beneficial effects on the gastrointestinal tract, growth performance, and nutrient absorption in pigs, often used as a probiotic in animal farming [58]. According to Gresse et al. [59], the prevalence of the Lactobacillus group declined, while bacteria like Clostridium, Prevotella, and facultative anaerobes, including Proteobacteriaceae, exhibited an overall increase in response. Prevotella excels at degrading high-fiber diets and is associated with enhanced intestinal immunity and reduced diarrhea [60]. This bacterium generates succinic and acetic acids through fermentation [61]. Ruminococcaceae are vital for breaking down complex carbohydrates and fibers, producing short-chain fatty acids that offer several benefits to the host, including energy provision, enhanced intestinal health, and support for immune system regulation [62]. Elevated levels of Ruminococcaceae are linked to reduced rates of constipation, inflammation, and liver diseases, which are commonly seen in individuals with better overall health [62,63]. The positive effects noted in our study can be ascribed to the probiotics’ role in promoting the growth of beneficial bacteria, which in turn enhances gut health and overall well-being in pigs. Furthermore, adding multi-strain probiotics altered the pig’s gut microbiota by enhancing the levels of potentially beneficial bacteria like Ruminococcaceae and declining the prevalence of pathogenic bacteria such as Escherichia and Chlamydia [20]. The presence of Christensenellaceae has been positively linked to feed efficiency, a pivotal aspect within the swine industry [64]. The incorporation of multispecies probiotics changed the activity of the gut microbiome, increasing pathways related to protein digestion and utilization, possibly resulting in enhanced production of metabolites within the gut [65]. Our study indicated an enhancement of the levels of Firmicutes, Prevotella, Lactobacillus, and a decline in Clostridium sensu stricto 1, Succinivibrio, and Proteobacteria in the treatment groups compared to the TRT1. The reason for these changes is likely due to the combined effects of ZnO and probiotics in promoting beneficial bacteria and suppressing pathogenic bacteria, thus creating a more favorable gut environment. Therefore, the biological data indicate that adding ZnO-probiotics to diets can help reduce gut infections, increase beneficial bacteria, and improve the immune system and overall health of pigs.
CONCLUSION
This study provides novel insights into the synergistic effects of combining lower levels of ZnO with probiotics in the diets of weaning-to-finishing pigs. The results demonstrate that this dietary strategy significantly enhances growth performance parameters, including BW, ADG, and ADFI, while simultaneously reducing noxious gas emissions (NH₃ and H₂S). Notably, our findings reveal that these improvements are closely associated with beneficial alterations in gut microbiota composition, characterized by an increased abundance of Firmicutes, Prevotella, and Lactobacillus and a reduction in harmful bacteria such as Clostridium sensu stricto 1 and Proteobacteria. Although no significant effects were observed on fecal scores or meat quality, the observed microbial shifts suggest a pivotal role of gut microbiome modulation in enhancing growth efficiency and mitigating environmental impact. This study contributes to the growing body of research on sustainable feeding strategies by demonstrating that reducing ZnO levels, when combined with probiotics, can effectively support pig health and performance while minimizing environmental pollution. Future studies should explore the long-term effects of this dietary strategy on immune function, intestinal integrity, and overall performance to further validate its benefits.