INTRODUCTION
Many microorganisms living in the intestine are known to play important roles in maintaining homeostasis and host health as commensal microorganisms by producing metabolites that cannot be synthesized in the body or inhibiting the growth of pathogens [1,2]. Among commensal bacteria, opportunistic pathogenic bacteria (pathobionts) exhibit pathogenicity depending on the environmental conditions [3,4]. Changes in the gut microbiome occur owing to various factors, such as the environment, feed, and pathogen infection. Among these, diarrhea caused by pathogen infection causes rapid changes in the gut microbiome [5–7]. Compared to healthy calves, the calves with diarrhea have reduced gut microbiome diversity and a microflora dominated by harmful bacteria [8]. In particular, newborn calves are easily infected with pathogens because their gut microbiome is not fully developed, and diarrhea caused by pathogen infection in the early stages of growth can have significant impacts on subsequent growth and can even lead to death [9,10].
McGuirk [11] defined diarrhea as feces with a fecal score of 2 (loose but consistent enough to remain on bedding) or 3 (watery feces that shift through bedding material). Diarrhea is a serious disease that affects newborn calves under 30 days of age and causes significant economic losses to farms [12–14]. Diarrhea in Hanwoo (Korean indigenous cattle) calves is caused by viruses (such as bovine viral diarrhea virus [BVDV], bovine rotavirus [BRV], bovine coronavirus [BCoV], etc.), protozoa (such as Giardia spp., Eimeria spp., Cryptosporidium spp.), and bacteria (such as Escherichia coli K99) [15–18]. Additionally, cases of simultaneous infection with two or more pathogens have been frequently confirmed [19]. In general, when diarrhea occurs in calves, clinical symptoms such as loss of appetite, dehydration, electrolyte imbalance, and metabolic acidosis appear. However, the infection mechanism differs depending on the type of pathogen, resulting in differences in clinical symptoms [17].
To date, many studies have been conducted on various pathogens causing diarrhea in Hanwoo calves and the resulting clinical symptoms; however, few studies have been conducted on the changes in the gut microbiome of Hanwoo calves following infection with each pathogen [20–22]. To prevent diarrheal symptoms caused by pathogen infection, vaccines, farm management, and colostrum intake are necessary. Early diagnosis of each pathogen using predictive indicators and accordingly treating and managing the calves are necessary. Confirming the relationship between pathogens and the gut microbiome of calves will help understand the effect of each pathogen on calves and will likely help prevent pathogenic infection. Therefore, the gut microbiomes of normal Hanwoo calves and those infected with BRV or BCoV with diarrheal symptoms were compared to analyze the effects of each pathogen on the gut microbiome.
MATERIALS AND METHODS
Fecal samples from eight Hanwoo calves under 30 days of age with diarrhea and seven of those without diarrhea were collected from several farms in Gyeongsangnam-do, Korea. The fecal consistency of diarrhea samples is loose or watery, while normal samples without diarrhea are solid and semi-solid. The collected feces were conducted pathogen tests and gut microbiomes were compared.
All fecal samples were collected directly from the calves’ anus by massaging the rectal wall with a finger to induce defecation. The collected feces were transported to the laboratory in a refrigerated state and stored at –20°C until analysis.
For BRV and BCoV testing in the collected fecal samples, 1 g of the fecal sample was placed in a 15 mL sterile tube (Conical Tube, SPL Life Sciences), mixed with 10 mL of PBS, and centrifuged at 3,000×g for 10 min. DNA/RNA was extracted from the supernatant using an automated extraction kit (AutoXT PGS DNA/RNA Kit; iNtRON). The extracted RNA was used to detect BVDV, BRV, and BCoV by real-time reverse transcription polymerase chain reaction (Real-time RT-PCR) using a commercialized bovine diarrhea virus triple test kit (PowerChek™ Bovine Disease Virus Triplex Real time PCR Kit, Kogene Biotech). The reaction solution was prepared by adding 5 μL of the extracted RNA to 15 μL of the Real-time RT-PCR Premix. Afterwards, the reaction was performed at 50°C for 30 min and 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min each [23].
The total genomic DNA of the microorganisms present in the 15 collected fecal samples was extracted using a DNA extraction kit (FastDNA SPIN Kit for Soil, MP Biomedicals). The V3/V4 region of the 16S rRNA gene was amplified by polymerase chain reaction (PCR) using the primers 341F and 805R. The metagenome of microorganisms extracted from each fecal sample was used as a template (PTC-200 Peltier thermal cycler, MJ Research; Table 1). The PCR conditions were pre-denaturation at 94°C for 3 min, followed by 28 cycles of denaturation at 94°C for 30 s, annealing at 53°C for 40 s, and elongation at 72°C for 1 min, followed by final extension at 72°C for 5 min. Afterwards, secondary amplification for attachment of the Illumina NexTera barcode was performed using the i5 forward primer and i7 reverse primer (Table 1). The PCR conditions were pre-denaturation at 94°C for 3 min, followed by 8 cycles of denaturation at 94°C for 30 s, annealing at 53°C for 40 s, and elongation at 72°C for 1 min, followed by final extension at 72°C for 5 min.
The amplified PCR products were purified using a QIAquick PCR Purification Kit (Qiagen) and subjected to electrophoresis to select DNA with a sequence length of 300 bp or longer. DNA fragment lengths were confirmed using an Agilent 2100 Bioanalyzer (Agilent Technologies). A library was constructed from the amplified products and sequenced using MiSeq (Illumina).
The base sequence data obtained through MiSeq results were classified by sample using the Mothur program (https://www.mothur.org). The paired-end reads for each sample were then made into a single contig, and sequence filtering was performed to meet the criteria through quality control [24]. The filtered reads were subjected to alpha diversity analysis (operational taxonomic unit [OTU], rarefaction curve, Shannon-Weaver, Chao1, etc.), and the microbial community structure and relationships at the phylum, class, order, family, genus, and species levels were identified using the EzBioCloud server (www.ezbiocloud. net/) and the CL community (ChunLab). Clustering was confirmed using the unweighted pair group method with arithmetic average (UPGMA), and beta diversity was measured using unweighted unique fraction metric (UNIFRAC) analysis [25–27]. Microbial changes were analyzed using principal coordinate analysis (PCoA) plots [28]. Linear discriminant analysis effect size (LEfSe) was used to identify bacterial taxa at p < 0.05, and a linear discriminant analysis (LDA) score > 2.0, using the Galaxy workflow framework (https://huttenhower.sph.harvard.edu/galaxy/).
The relative abundances of major phyla, classes, orders, families (median relative abundance > 0.1%), and major genera (median relative abundance > 0.01%) were calculated, and comparisons between the groups were performed using GraphPad Prism 6 (GraphPad Software). Normality was analyzed using the Shapiro-Wilk test, and comparisons between groups were performed using the Kruskal-Wallis test. In all statistical analyses, significance was set at p < 0.05.
RESULTS
Of the eight calves with diarrhea, BRV was detected in four and BCoV was detected in four, all of which were confirmed to be single infections rather than multiple infections. No pathogens were detected in the normal calves without diarrhea.
To compare the differences in the gut microbiome based on the cause of infection in Hanwoo calves, 864,232 sequence reads (an average of 57,6158 reads per calf) were obtained from the feces of four calves with BRV, four with BCoV, and seven normal calves, securing a sufficient number of OTUs required for analysis.
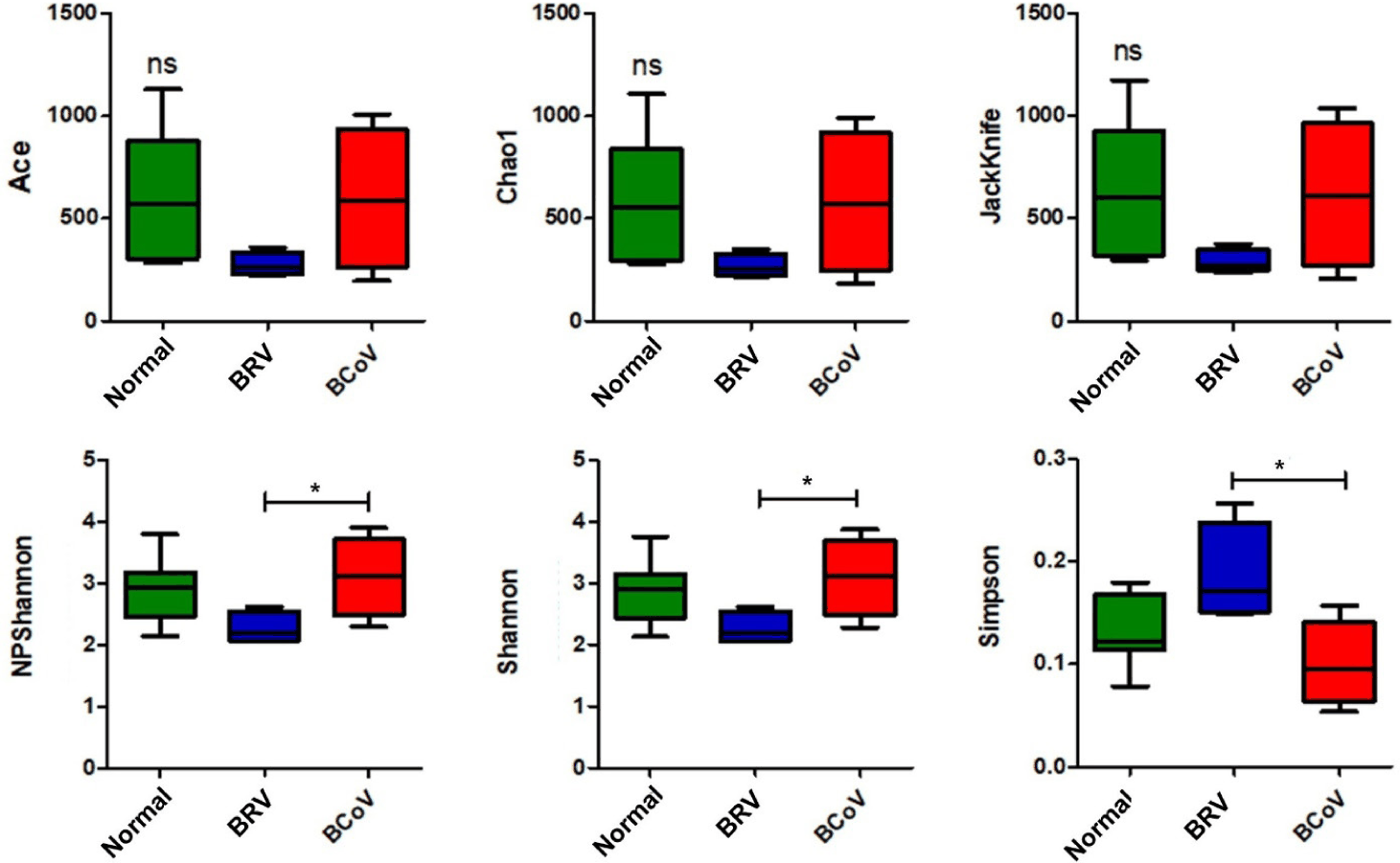
Alpha diversity analysis was performed on richness and evenness indices to determine the species diversity and distribution of the gut microbiome. There were no significant differences in the richness and evenness indices among the normal, BRV-infected, and BCoV-infected calves. However, there were significant differences in the NPShannon, Shannon, and Simpson evenness indices between calves with BRV and those with BCoV (Fig. 1).
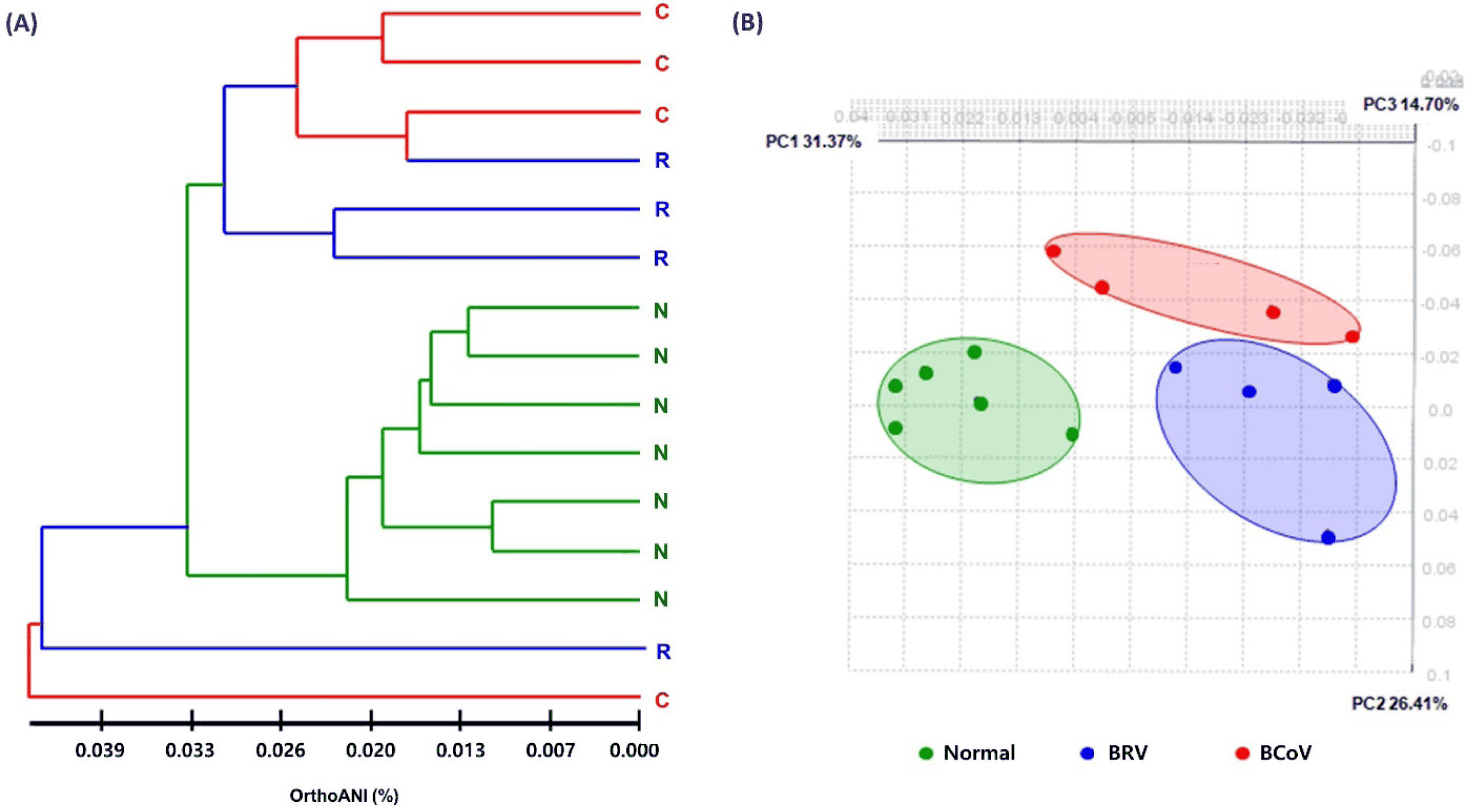
Clustering and beta diversity analyses were performed to compare and analyze the species diversity of the gut microbiome and confirm the similarity relationship. Clustering showed that the microbiome were clearly differentiated between normal and BRV-infected calves, and between normal and BCoV-infected calves, and this was consistent with the results of PCoA analysis (Fig. 2).
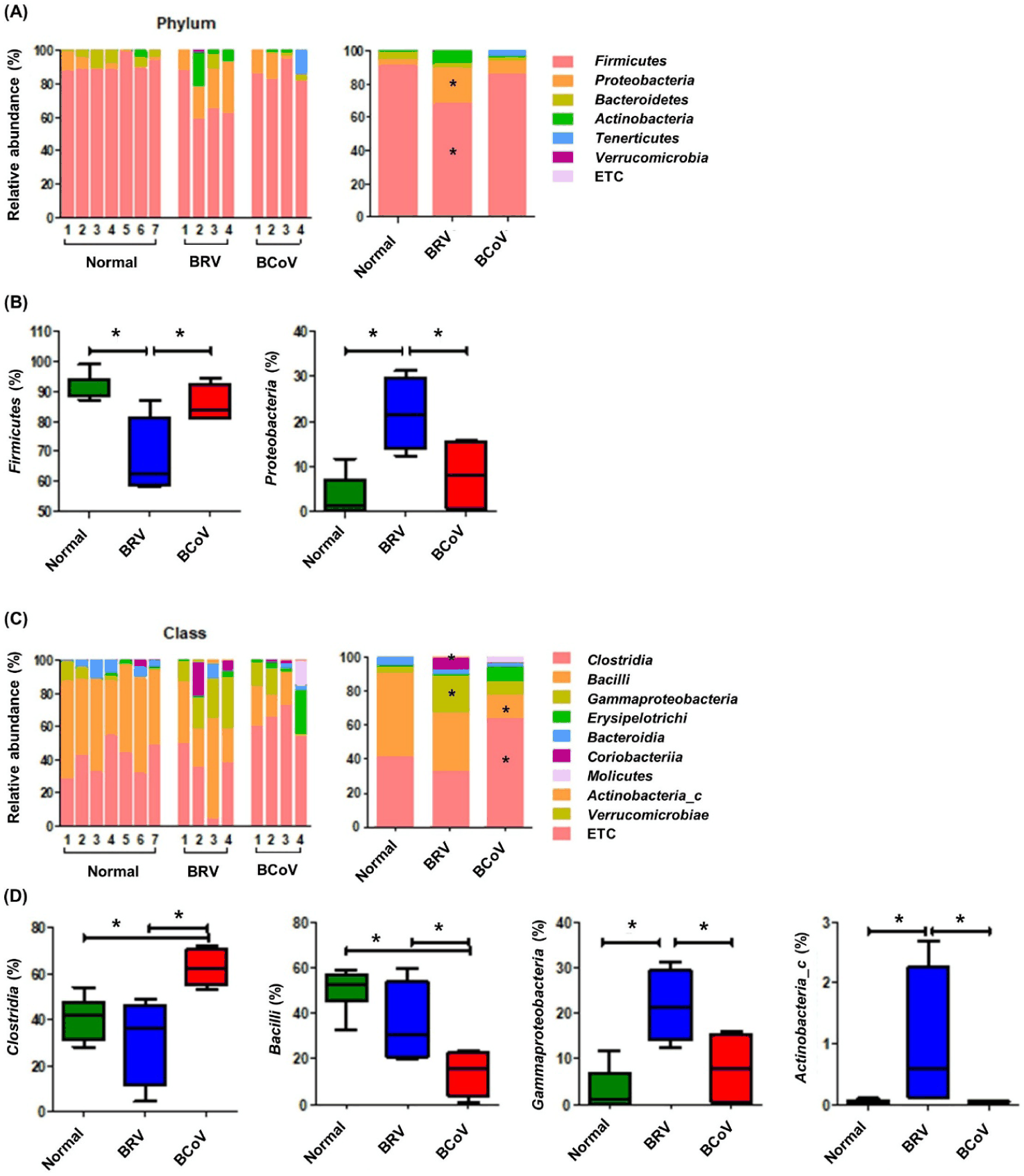
A comparison of the gut microbiomes of normal calves, those infected with BRV, and those with BCoV at the phylum level showed that calves infected with BRV showed a decrease in Firmicutes and an increase in Proteobacteria, which were significantly different from those of normal calves and those infected with BCoV (Fig. 3A). Compared with normal calves, those infected with BRV showed a significant increase in Gammaproteobacteria and Actinobacteria at the class level, and those infected with BCoV showed a significant increase in Clostridia and a decrease in Bacilli (Fig. 3B).
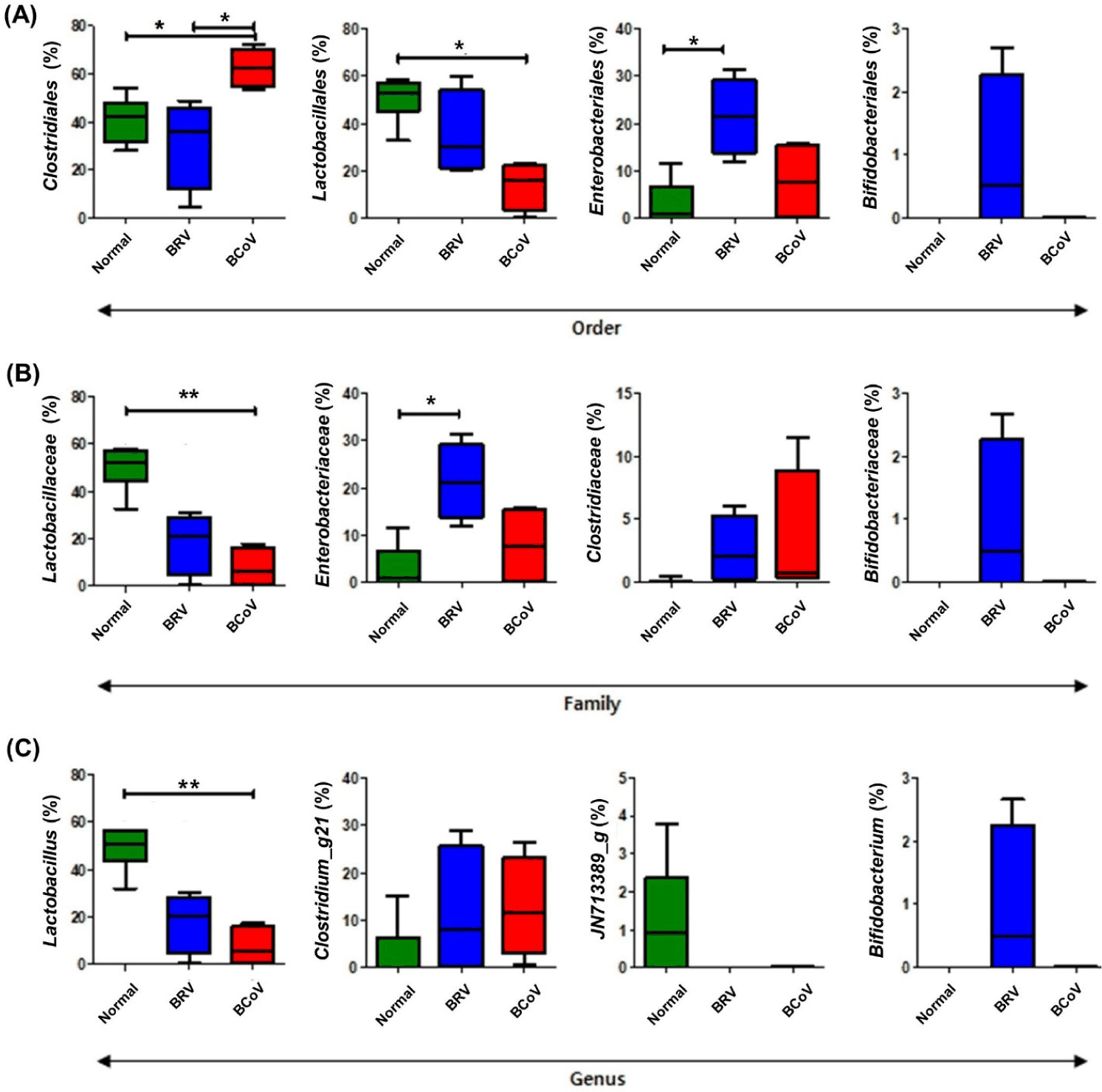
At the order level, calves infected with BRV showed a significant increase in Enterobacteriales, whereas those infected with BCoV showed a significant increase in Clostridiales and a significant decrease in Lactobacillales (Fig. 4A). At the family level, calves infected with BRV showed a significant increase in Enterobacteriaceae (Fig. 4B). At the genus level, Bifidobacterium was increased in rotavirus-infected calves, although not significantly (Fig. 4C). In addition, the relative frequencies of Lactobacillaceae and Lactobacillus significant decreased in both BRV- and BCoV-infected calves at the family and the genus levels, respectively (Figs. 4B and 4C).
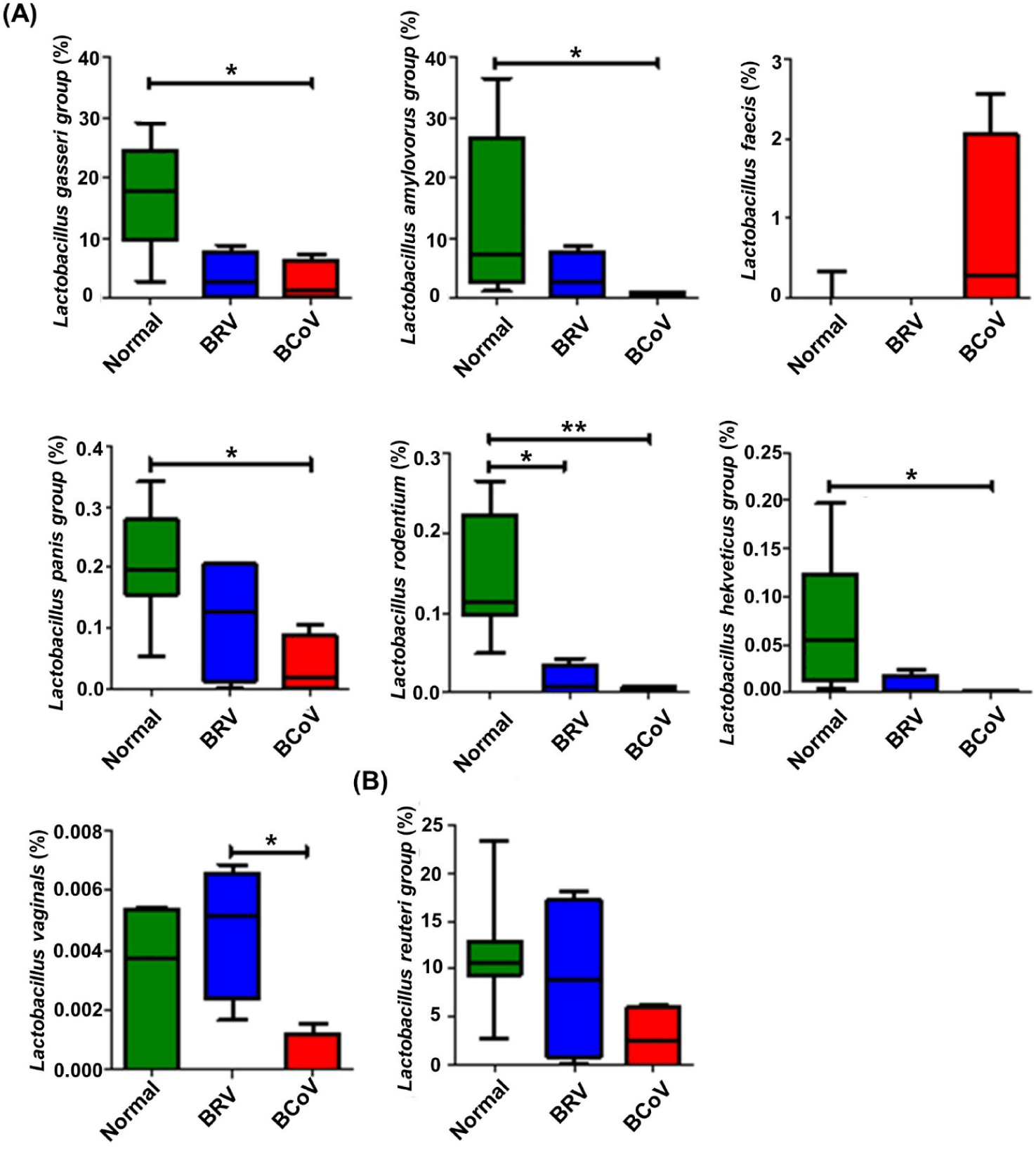
The gut microbiota associated with normal, BRV-infected, and BCoV-infected calves were identified through LEfSe analysis. As a result, 2 phyla, 4 classes, 4 orders, 4 families, 4 genera and 16 species were identified (Fig. 5A). The results of the cladogram based on LEfSe analysis showed that normal calves and those infected with BRV showed differences in the distribution of Firmicutes and Proteobacteria at the phylum level, whereas normal calves and those infected with BCoV showed differences in Clostridia and Bacilli at the class level (Fig. 5B and 5C).
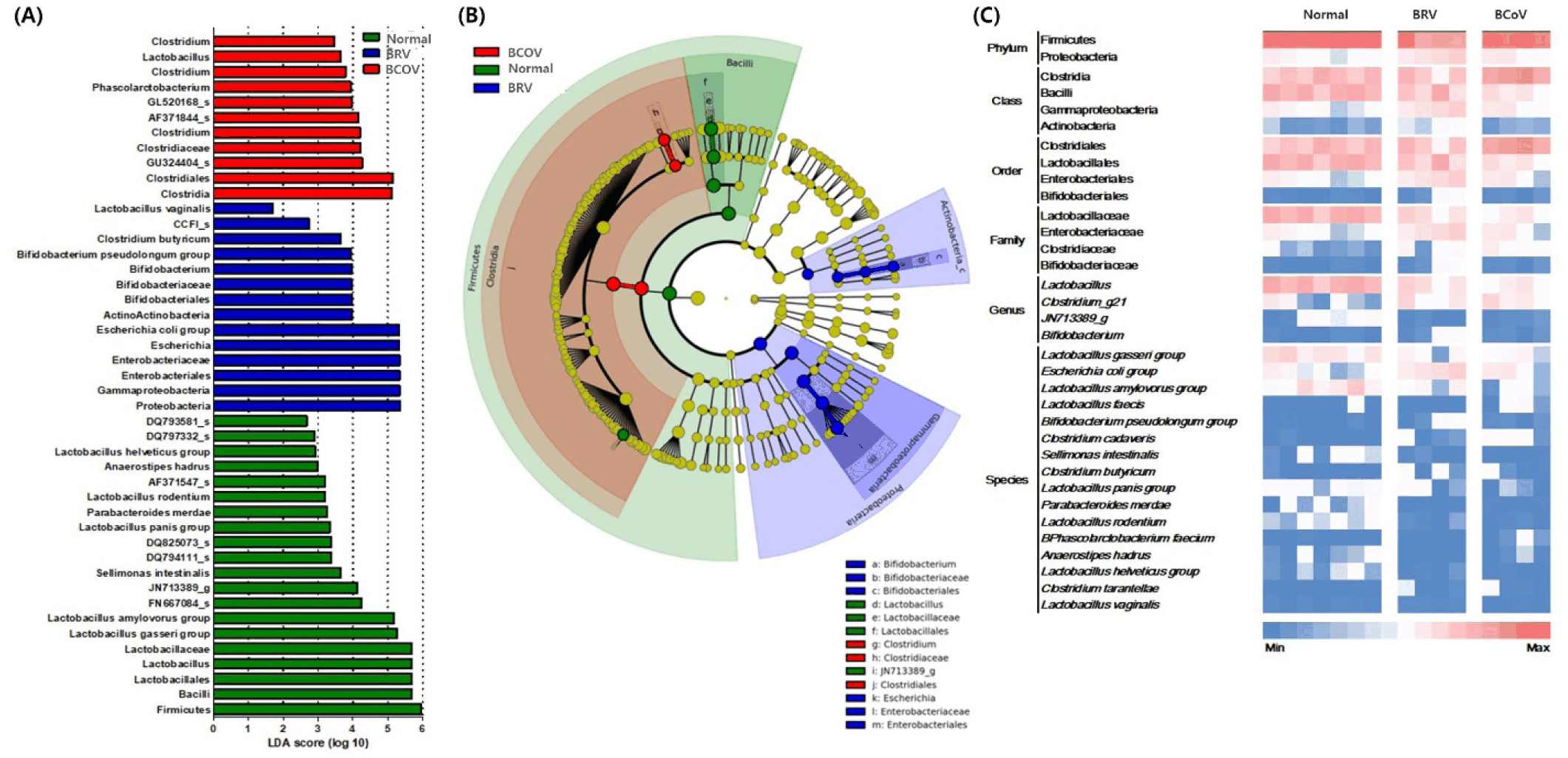
The relative abundance of seven species of Lactobacillus with LDA values higher than 2.0 in normal calves were analyzed, and the Lactobacillus gasseri, Lactobacillus amylovorus, Lactobacillus panis, Lactobacillus helveticus groups; Lactobacillus rodentium; and Lactobacillus vaginalis tended to decrease with BRV or BCoV infection. Specifically, L. rodentium group were significantly decrease in BRV or BCoV infection calves. In addition, L. gasseri, L. amylovorus, L. panis and L. helveticus groups showed a significant decrease in BCoV infection calves compared to normal calves. However, Lactobacillus faecis increased only in the calves infected with BCoV (Fig. 6A and Supplementary Fig. S1). Although there was no significant difference between the groups, the relative abundance of L. reuteri was more than 10% (Fig. 6B).
DISCUSSION
To investigate the differences in the gut microbiome of Hanwoo calves based on the pathogen causing diarrhea, calves with diarrhea were selected and infection with BRV or BCoV was confirmed. Comparison of the gut microbiomes of these calves revealed distinct differences between normal calves and those infected with BRV and BCoV. Changes in the taxonomic composition were also observed.
Diarrhea reduces gut microbiome diversity [29]. In this study, the number and diversity of gut microorganisms were reduced in BRV-infected calves compared to normal calves, but there was no difference in BCoV-infected calves. However, a previous study reported that the number of gut microbial species increased in calves infected with BRV, and that both BRV and BCoV infections reduced gut microbiome diversity [30]. In addition, this study showed that the microbiome was distinctly different among the normal calves and those infected with BRV and BCoV. These differences in results might have occurred because various factors, such as the environment, feed, and age of the calf, as well as the time elapsed after pathogen infection, affect the gut microbiome. In addition, because the number of calves tested was limited, more research is required with larger sample sizes.
At the phylum level, the gut microbiome of calves is dominated by Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria [7,31]. In this study, the relative frequencies of Firmicutes and Bacteroidetes decreased, whereas those of Proteobacteria increased in the gut microbiome of calves infected with BRV compared to normal calves. This change was confirmed to be similar to that of the gut microbiome of BRV-infected calves reported previously [30,32]. However, the gut microbiome of calves infected with BCoV was not significantly different from that of normal calves. It is known that in calves with diarrhea, Firmicutes and Proteobacteria increase, while Bacteroidetes decreases, resulting in an increase in the Firmicutes/Bacteroidetes ratio [33]. Therefore, a decrease in Firmicutes in the gut microbiome of calves is thought to be a characteristic of BRV infection.
There are an increasing number of reports on changes in the gut microbiome in response to pathogens that cause diarrhea. Cryptosporidium spp., Eimeria spp., and Giardia spp., which are frequently detected in Hanwoo calves with diarrhea, have also been found to cause imbalances in the gut microbiome during infection [34–36]. However, studies targeting pathogens in calves are limited, and only a few have been conducted on Hanwoo calves. In addition, in this study, the results of the cladogram based on LEfSe analysis showed that normal calves and those infected with BRV showed differences in the distribution of Firmicutes and Proteobacteria at the phylum level, whereas those infected with BCoV showed differences in Clostridia and Bacilli at the class level. This is similar to previous studies that showed significant differences in the gut microbiome depending on the pathogen causing diarrhea in calves, and it is necessary to select specific microorganisms with a correlation [22,30,37]. In the present study, protozoan parasites (Cryptosporidium spp., Eimeria spp., and Giardia spp.) were not detected in the feces of the selected calves (data not shown), and only BRV and BCoV were identified. However, because infections with other potential pathogens may exist and influence the results, further studies targeting various pathogens are expected to elucidate the association between BRV and BCoV and the gut microbiome, which will help predict and diagnose diarrhea in calves.
Bifidobacterium is the dominant species in the intestine of newborn calves and is found in high relative abundance in healthy calves [38]. Bifidobacterium produces lactic acid and short-chain fatty acids (SCFAs) and can inhibit the colonization of pathogenic bacteria in the intestines, so it is important for the health of calves, and Bifidobacterium strains are used as probiotics [39–41]. However, Bifidobacterium was hardly present in the fecal of most calves in this study. Although the relative abundance was high in some BRV-infected calves, the difference between individuals was large and there was no significant difference compared to other groups. Although the cause is still unknown, it has been reported that Bifidobacterium in the gut microbiome of calves are at their highest on the 7th day after birth and then decrease with growth [42–44]. In addition, the calves used in this study were at a stage where they consume a mixture of milk and concentrate feed, which is thought to have affected the decrease in Bifidobacterium [45]. In addition, calves infected with BRV and BCoV showed a decrease in Lactobacillus compared to normal calves. In calves infected with BRV, where Lactobacillus was significantly reduced, Bifidobacterium was significantly increased, whereas in calves infected with BCoV, Lactobacillus was significantly reduced and Clostridium g21 was increased. This suggests that the composition of the intestinal microbiota changes depending on the pathogen. However, previous studies have reported an increase in Lactobacillus in calves with diarrhea [8,32,46,47]. When calves show diarrhea symptoms, D-lactate and L-lactate levels increase, which increases lactic acid-producing bacteria such as Lactobacillus, and the decrease in intestinal pH aids the growth of acid-stable Lactobacillus. However, a previous study reported that Lactobacillus abundance decreases 24 h before the clinical manifestation of diarrhea [48]. Therefore, in this study, it is possible that the time elapsed between the onset of diarrhea and sampling and the clinical condition of the calves caused the difference in the Lactobacillus ratio. In addition, because Lactobacillus is expected to be related to the health status of Hanwoo calves, Lactobacillus can be utilized as a useful microorganism for maintaining the intestinal environment of healthy Hanwoo calves. In particular, unlike calves infected with BRV, the calves infected with BCoV showed increase in Lactobacillus faecis. Hence, L. faecis can be used as an indicator for the prevention and diagnosis of BCoV in calves through gut microbiome analysis. In addition, Clostridium, Enterococcus, and Escherichia, which are significantly associated with diarrhea in calves, are the main microorganisms that play a pathogenic role, and it is thought that these microorganisms have a growth advantage in the changed intestinal environment caused by diarrhea [49]. Therefore, diarrhea caused by BRV and BCoV infections can make calves more vulnerable to pathogenic microorganisms, worsen the imbalance of the gut microbiome, and decrease their immune function.
BRV and BCoV, the main pathogens causing diarrhea in calves, showed significant differences in the gut microbiome compared to that in healthy calves that did not show diarrhea when infected. In addition, significant differences in the gut microbiome were confirmed, depending on the pathogen. Analysis of the gut microbiome targeting each pathogen, which has rarely been studied so far, has revealed microorganisms associated with each pathogen, and utilizing these microorganisms as indicators can help improve the early detection of diseases and treatment efficiency through standardized physiological indicators.