
INTRODUCTION
Milk production by dairy cows has increased by 41% (2,802 kg/head) in milk yield and 0.12% (120.04 kg/head) in milk fat from 2,000 to 2,020 in Korea [1]. With these improvements and selection, dairy cows are more sensitive to heat stress, which directly or indirectly influences the normal physiology, metabolism, and hormonal and immune systems [2]. Heat stress also causes economic losses according to negative effects on the milk production, milk composition, reproductive efficiency, and udder health [2,3]. Thus, environmental and nutritional methods to alleviate heat stress have been studied to improve the lactation performance of dairy cows.
In particular, feed additives, including yeast supplements such as Saccharomyces cerevisiae, are widely used to improve the lactation performance and health in dairy cows [4,5]. Many studies have reported that feeding S. cerevisiae positively influences the dry matter intake (DMI), rumen pH, nutrient digestibility, immune function and milk quality of dairy cows [5–7]. Because of its effects on the rumen fermentation and nutrient digestion, some authors have suggested that feeding yeast products may be most beneficial to dairy cows under heat stress [8–12]. Arambel and Kent [8] suggested that yeast products might be more effective under heat stress than under normal conditions. Schingoethe et al. [9] reported a significant improvement in feed efficiency when mid-lactating dairy cows were supplemented with S. cerevisiae during summer months. S. cerevisiae culture contains fermentation substrates, bacterial proteins, yeast metabolites, yeast cell walls, and other beneficial substances, and it can balance the interstinal flora, improve immunity, relieve stress, and improve productivity [10].
Recently, several agricultural technology centers in Korea have supplied microbial cultures such as Lactobacillus plantarum, Bacillus subtilis, and S. cerevisiae, which are known to effectively improve the feed efficiency, productivity and environment of livestock. However, research on the method or effectiveness of feeding S. cerevisiae culture to dairy cows is limited. There is also limited understanding of how S. cerevisiae culture affects the lactation performance of heat-stressed dairy cows associated with blood metabolism. Therefore, we hypothesized that S. cerevisiae culture would improve the milk productivity and blood metabolism of dairy cows exposed to heat stress if feeding S. cerevisiae could increase the DMI of dairy cows under such conditions. To address this hypothesis, the effects of the S. cerevisiae culture addition in different total mixed rations (TMRs) on DMI, milk production and quality, and energy status in dairy cows during heat stress were evaluated.
MATERIALS AND METHODS
All dairy cows and the experimental protocol in this experiment were approved by the Institutional Animal Care and Use Committee (IACUC, study approval number: 20191519) at the National Institute of Animal Science (NIAS, Jeonju, Korea). The experiment was conducted at the Department of Animal Resources Development, NIAS (Cheonan, Korea). Twenty-four Holstein dairy cows were allocated into 4 groups based on day in milk (DIM) (187.3 ± 41.9; mean ± standard deviation), parity (1.75 ± 0.19), milk yield (31.21 ± 1.05 kg/d), and body weight (BW) 678 ± 10.67 kg. The cows were randomly assigned within blocks to one of four dietary treatments: (Control) TMR-1; (T1) TMR-1 supplemented with S. cerevisiae culture fluid (SCCF, 5 mL/head); (T2) TMR-2; and (T3) TMR-2 supplemented with SCCF (Table 1). TMR-1 was mixed with domestic forages such as corn silage, rice straw silage, and whole-crop barley silage. TMR-2 was mixed by adding alfalfa hay, which has mainly been used in dairy farms, in addition to domestic forages of TMR-1. SCCF (S. cerevisiae, ca. 2.0×107 CFU/mL, AAH180004, Anseong 40-18-83-100), which is a fully fermented yeast culture containing fermentation metabolites, residual yeast cells, and growth media, was provided by the Agricultural Technology Center (Anseong, Korea). The SCCF supplementations were mixed with TMRs daily at the time of feeding to dairy cows. TMR-1 and TMR-2 were formulated to satisfy or exceed the nutrient requirements [13] for lactating Holstein dairy cows (DMI 22.0 kg/d, net energy for lactation [NEL] 32.5 Mcal/d). The ingredient and nutrient contents of the feed components are presented in Table 1.
TMR, total mixed ration; SCCF, Saccharomyces cerevisiae culture fluid (Agriculture Technology Center, Anseong, Korea); DM, dry matter; DDGS, dried distiller’s grains with solubles; NDF, neutral detergent fiber; ADF, acid detergent fiber; NEL, net energy for lactation; RFV, relative feed value; RFQ, relative feed quality
Cows were housed in a loose barn and fed the TMR daily at 09:00 with free access to drinking water. Feed was offered ad libitum to yield 10% residues. The barn contained fans over feeding alleys every 5 m, and the fans were operated once the ambient temperature reached 25°C. The experiment commenced from July 12 to August 8, which is the typical hot season in Cheonan, Korea. The feeding trial was composed of a 2-week adaptation period and a 2-week experimental period.
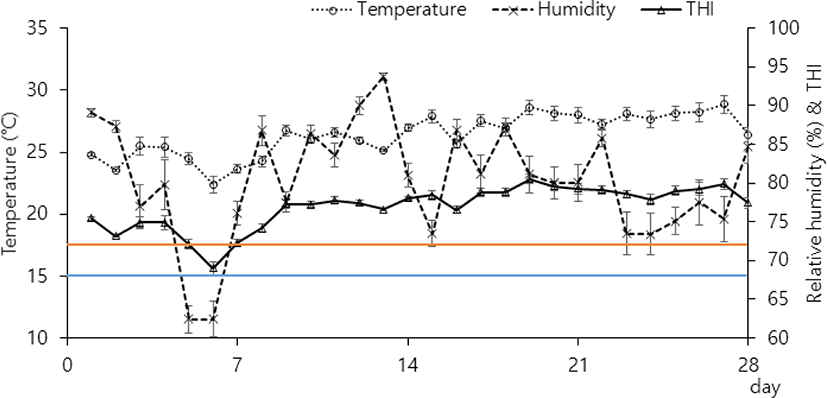
To measure the environmental conditions inside the barn, temperature and relative humidity (RH) were recorded by a thermohygrometer (Model 174H, Testo, West Chester, PA, USA) with an accuracy of ± 0.5°C and ± 3% RH. A thermohygrometer was set to record every hour per day and placed at a height of 2 m from the feeding area. The temperature-humidity index (THI) was calculated as THI = (0.8 × C) + [RH % × (C − 14.4)] + 46.4 according to Mader et al. [14]. For each 24 h period, the average daily minimum, mean and maximum temperatures and RH were determined using the hourly recorded data, and the minimum, mean and maximum THIs were calculated (Fig. 1 and Table 3). Feed offered and residues were weighed daily to determine the DMI throughout the experiment. Representative samples of individual dietary ingredients and TMR were collected weekly and stored at –20°C until analyses. All samples were analyzed by AOAC [15] for concentrations of dry matter, crude protein, ether extract, crude fiber, and crude ash and by Van Soest et al. [16] for concentrations of neutral detergent fiber (NDF) and acid detergent fiber (ADF). Particle size distributions of TMR-1 and TMR-2 were measured by sieving using the Penn State Particle Separator [17]. The sum of the DM retained on the top (19 mm) and middle sieves (8 mm) was assumed to be equal to the physical effectiveness factor (pef) of feeds.
Cows were milked 2 times daily at 06:00 and 17:00 h, and individual milk production was recorded daily using a DeLaval’s Alpro Herd management system (DeLaval international, Tumba, Sweden) with a double-eight herringbone parlor. Individual milk samples were collected from consecutive morning and afternoon milking on the seventh day of each week from each cow. The milk samples were analyzed to obtain the somatic cell count (SCC), fat, protein, lactose, total solids, and milk urea nitrogen (MUN) concentrations using a CombiScope FTIR (Delta Instruments, Drachten, Netherlands).
All cows were sampled for blood on the last day of the study. Blood samples of each cow were taken through the jugular vein with a syringe before feeding the cows. The collected blood samples were transferred into vacutainer tubes (BD Vacutainer, Becton Dickinson, Frankin Lakes, NJ, USA) and centrifuged at 1,000×g for 15 min at 4°C. The serum samples were immediately frozen at -20°C and later analyzed for aspartate aminotransferase (AST), alanine aminotransferase (ALT), glucose, triglyceride, cholesterol, nonessential fatty acid (NEFA), blood urea nitrogen (BUN), and creatinine concentrations. Serum samples were used to analyze biochemistry (Wako Chemicals, Neuss, Germany) using a blood autoanalyzer (Hitachi 7180, Hitachi, Tokyo, Japan).
Statistical analysis was conducted using the Statistical Analysis System (SAS) Enterprise Guide 7.1 (SAS Institute, Cary, NC, USA). The particle size distribution was analyzed to determine the effects of the experimental TMRs with one-way analysis of variance (ANOVA). The DMI, BW gain, milk yield, milk composition, and blood metabolite data were analyzed by two-way ANOVA using the general linear model procedure to determine the main effects, treatments (with or without SCCF) and TMR (no vs. mixing the alfalfa hay), and their interaction (Treatment × TMR). If there were differences, we continued with the Duncan multiple range test. Differences with p < 0.05 were declared significant. Duncan’s multiple range test was used to evaluate the significant differences.
RESULTS AND DISCUSSION
The average mean daily temperature (°C) and RH (%) during the entire experiment were 26.40 ± 0.32°C and 80.32 ± 1.39%, respectively (Fig. 1). The average mean daily THI (76.92 ± 0.51) in the barn housing cows over all days exceeded 68, which characterizes exposure to heat stress for lactating dairy cows [18]. The daily THI variation was 8.14 ± 0.54 on average and ranged from 73.04 to 81.19 (Table 2).
A THI value of 72 is a critical threshold value, above which the productive properties of the cow begin to change and a drop in productivity is noted [19]. Recent studies state that THI 68 is the lower limit for the occurrence of heat stress [18]. Carter et al. [18] noted that high-yield dairy cows experienced heat stress when average daily THI > 68 or minimum daily THI > 65. From previous studies, environmental conditions over all days caused heat stress for lactating dairy cows.
Table 3 shows that TMR-2 had a lower proportion retained on the top screen (> 19 mm) and a higher proportion retained on the middle screen (8–19 mm) than TMR-1 (p < 0.05). The particle size distribution of TMR-2, which replaced some of the long forage such as whole crop barle (WCB) and rice silage with alfalfa hay, resulted in lower physically effective neutral detergent fiber (peNDF)19.0 and peNDF8.0 than those in TMR-1 (p < 0.05).
The particle size distribution of TMRs in this study did not satisfy the recommendation guidelines suggested by Heinrichs and Kononoff [20]: 2%–8% long particles (> 19 mm), 30%–50% medium and short particles (8–19 mm), and no more than 20% fine particles (< 8 mm). A particle fraction > 19 mm and peNDF8.0 were good parameters for monitoring the TMR physical form indicator to improve the chewing activity and rumen pH and prevent sorting [21]. In this study, beneficial physical effects were not observed for TMR with added alfalfa hay because TMR-2 had fewer particles > 19 mm but more particles < 8 mm than TMR-1.
The intake of DM in all treatments did not differ by the type of TMR (TMR-1 and TMR-2) (Table 4). This result is not consistent with Fischer et al. [22], who observed that the DMI was higher for those fed hay and TMR containing short instead of long silage. Meanwhile, the DMI was 2.7 kg/d higher for T1- and T3-supplemented S. cerevisiae than for the control and T2 cows (p < 0.05). Similar tendencies were observed in some studies: the DMI in dairy cows was affected by S. cerevisiae supplementation during early lactation [23] or mid-lactation during the summer [11]. Heat-stressed cows reduce the feed intake, rumination and saliva production. It may act as a risk factor to make the rumen function inappropriate [11]. S. cerevisiae can alter the ruminal microbial fermentation by stimulating the activity of fiber-digesting bacteria and increasing fiber digestion [6]. From the present study, feeding S. cerevisiae may mitigate the negative effect of heat stress on the DMI of dairy cows.
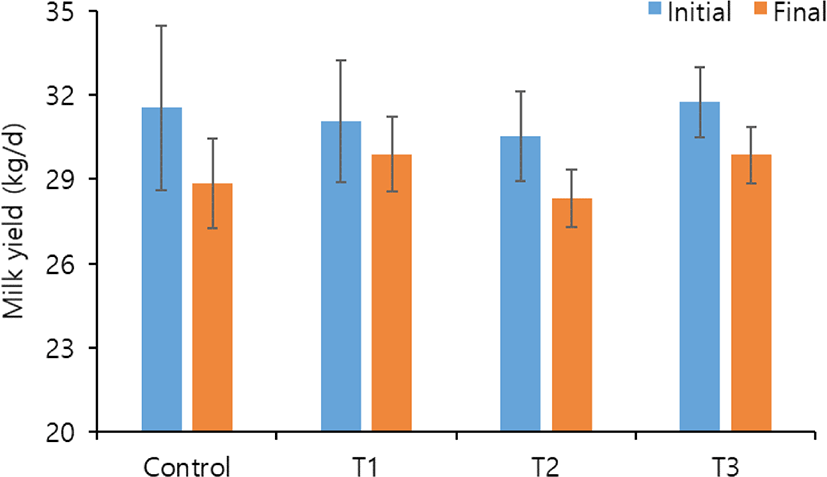
Milk yield were not affected among the treatments. However, some studies reported that the milk yield increased with increasing levels of S. cerevisiae under normal conditions [11]. Additionally, when heat-stressed dairy cows were fed yeasts and yeast cultures, milk production increased by 1–2 kg/d [23,24]. In this study, the reduction of milk yield numerically was lower in T1 and T3 (−1.16 and −1.89 kg/d, respectively) with supplemented S. cerevisiae than in control and T2 (−2.69 and −2.23 kg/d, respectively) with prolonged exposure to heat stress compared with the beginning of this trial (Fig. 2). Dairy cows exposed to heat stress exhibited decreased milk yield due to the high metabolic heat production with ruminal fermentation [2,3]. However, according to Callaway and Martin [6], the beneficial effects of S. cerevisiae supplementation on milk production resulted from the stabilized condition of rumen fermentation.
The feeding treatment of S. cerevisiae had no impact on the concentration of fat, protein, and lactose. These results are consistent with Arambel and Kent [8], who found that S. cerevisiae supplementation had no positive effect on the milk composition of dairy cows. Specifically, no changes in percentage of milk fat and protein appeared in lactating dairy cows under heat stress [9,11]. However, the total solids in milk were higher in dairy cows that were fed SCCF than in those not fed SCCF (p < 0.05).
SCC is a commonly used index of milk quality and a defense component of mammary glands against disease or intramammary infections [25]. The SCC in the present study was reduced by feeding S. cerevisiae (p < 0.05). Similarly, some studies reported that yeast product supplementation improved the mammary gland health in lactating dairy cows and reduced the incidence of mastitis [4,7]. These results are not fully explained but may be due to immunomodulation with antioxidant and anti-inflammatory effects of yeast product supplementation [26]. Although it is difficult to clearly explain the relationship between S. cerevisiae supplementation and SCC in milk from this study, yeast feed addition appears to help alleviate the increase in SCC during heat stress in a previous study.
Feeding SCCF decreased the MUN concentration in milk (p < 0.05), which corroborates previous studies [11] indicating that S. cerevisiae supplements reduced MUN during natural heat stress. Other studies observed that heat stress increased MUNs, and feeding SCCFs had no detectable effects on MUNs [12]. Heat-stressed cows mobilize skeletal muscle to provide amino acids to produce acute-phase proteins [27] and glucose [28]. The difference in the results of previous studies and this study may be due to different stages of lactation, feeding strategy (type of forage fed, feeding strategy, and forage-to-concentrate ratio [5, 29]), and heat stress severity [12].
The NEFA concentrations were slightly different between treatments with or without SCCF supplementation and, according to control, BUN was affected on the TMR groups either with alfalfa hay or not (p < 0.05; Table 5). The other metabolic indices, including AST, ALT, glucose, triglyceride, cholesterol, and creatinine, were not affected by the SCCF supplementation or experimental TMRs.
Some studies observed that the blood parameters were not affected by the S. cerevisiae treatment [12,29]. However, other studies reported that S. cerevisiae supplementation favorably influenced the energy metabolic status and might have a liver-protecting effect on high-yield cows [30,31].
Cows in a state of negative energy balance are biochemically characterized by elevations in blood biochemical parameters such as β-hydroxybutyrate (BHB) and NEFA and reductions in glucose concentrations [32]. In the current study, a lower NEFA concentration was observed in dairy cows fed SCCF (p < 0.05). This result is consistent with the results reported by Kumprechtova et al. [30] that the NEFA level was lower in early lactation dairy cows fed S. cerevisiae supplementation. Their study explained that lower NEFA levels might be considered a sign of reduced lipomobilization processes and hepatic ketogenesis. Dairy cows that were offered TMR with alfalfa had lower BUN concentrations, probably as a result of the lower total N and energy consumption than those of TMR with whole-crop barley.
The effects of SCCF on the lactation performance and blood metabolism of dairy cows remain inconsistent among previous studies. However, little research has been conducted on the effects of S. cerevisiae dietary supplementation on heat-stressed dairy cows, so further study relating to this issue is required.
CONCLUSION
In the present study, SCCF supplementation was fed to mid-lactating dairy cows during heat stress under the conditions with different ingredients of TMR to improve the milk productivity and blood metabolism. The findings of this study reveal that S. cerevisiae alleviated the reduction in DMI and the increase in SCC in heat-stressed dairy cows but did not affect the milk composition. Feeding S. cerevisiae contributed to a decrease in NEFA concentration, which reflects the energy metabolism, but did not affect the other blood parameters. Thus, SCCF supplementation can increase the DMI and improve the milk quality.