
INTRODUCTION
The formation of the biological microenvironment for sperm maturation and storage in the epididymis occurs via various specific events. One of the most important events is cell–cell communication among the principal, basal, and clear cells of the epididymal epithelium to create a specialized environment [1-3]. In addition, the blood–epididymis barrier (BEB) is comprised of tight junctions (TJs) between neighboring epithelial cells, which also supports the creation and maintenance of proper physiological conditions for the maturation and storage of spermatozoa [4-6]. In male reproductive tracts, the distribution of various tight junctional proteins is well reported in the testis and the epididymis, especially. In the testis, the blood–testis barrier (BTB) is located close to the base of the seminiferous tubule between adjacent Sertoli cells [7,8]. In the epididymis, however, the BEB is formed at apical surfaces of the epididymal duct between epithelial cells [9,10]. The BTB and BEB show positional differences but have the same function, which is to regulate the movement of electrolytes, ions, water, nutrients, hormones, and molecules, and the permeability of the paracellular transport pathway. The BTB and BEB also maintain the immunologically privileged adluminal compartment, which is a necessary component of the functioning barrier of the testis and epididymis [11-16].
TJs are protein complexes composed of several peripheral membranes and integral transmembrane proteins. Zonula occludens-1 (ZO-1) was first discovered as a TJ component in epithelia located on the intracellular side of the plasma membrane [17]. Additionally, two more proteins, ZO-2 and ZO-3, were confirmed as members of the ZO protein family [18,19]. These ZO proteins play a critical role in forming bridges between trans-membrane associated proteins and the actin cytoskeleton [20,21]. In contrast, occludin was first confirmed as the integral transmembrane protein and has four transmembrane domains with intracellular N- and C-termini [22]. In addition, claudins (Cldns) are also known as TJ transmembrane proteins, comprising at least 27 members in mammals [23,24]. Both occludin and Cldns interact with various intracellular proteins of TJs, including ZO-1, ZO-2, and ZO-3 [25,26]. TJ complexes that form a variety of proteins can create a physical, physiological, and immunological barrier.
Recent studies have reported that hormones can regulate the expression levels and localization patterns of TJ proteins in the testis. For example, gonadotropin hormones (GH), follicle stimulating hormone (FSH), and luteinizing hormone (LH) regulate the formation of the pituitary gland and are necessary for TJ formation in rat testes because GH plays a critical role in the development and differentiation of Sertoli cells where TJs are formed [27,28]. Androgens can upregulate Cldn3 and Cldn11 in Sertoli cells [13,29], and the administration of flutamide, an androgen receptor antagonist, disrupts Cldn11 protein in rat testes [30]. Additionally, in the epididymis, we reported that the extracellular signal-regulated kinase (ERK) signaling pathway is a key player in controlling the expression and localization of TJ proteins, such as ZO-2, ZO-3, occludin, and Cldn1, 3, and 4 in TJs of the mouse epididymis [31]; however, we know less about the functional roles of TJs in the epididymis than in the testis. Moreover, there has been no study on the expression and localization of TJ proteins and changes in TJ proteins during postnatal development of the epididymis in goats. Hence, in this present study, we characterized the distribution and localization patterns of TJ proteins in the immature and mature epididymides of goats using immunofluorescence and western blotting analyses.
MATERIALS AND METHODS
All information of primary and secondary antibodies used for immunofluorescence staining and western blotting is described in Tables 1 and 2, respectively.
Epididymides were collected using a surgical castration procedure from immature (3-month-old, n = 4), and mature (14-month-old, n = 5) black goats with an average body weight of 9.95 ± 0.66 kg and 28.04 ± 1.42 kg, respectively, at the National Institute of Animal Science. All animal experiments used here were performed under regulation and permission of the Institutional Animal Care and Use Committee (IACUC) of the National Institute of Animal Science (NIAS, Approval No. NIAS 2018-290).
Following castration, the epididymides were briefly cleaned in a cold phosphate-buffered saline (PBS) buffer to remove blood. The fixation of the epididymides was achieved by immersion in 4% paraformaldehyde solution (PFA) for 48–72 hr at 4°C, and the fixative was replaced with fresh solution every 24 hr. The samples were rinsed with PBS five times for 30 minutes each at room temperature (RT). To prevent ice crystal formation during freezing tissue samples, the epididymal tissues were immersed into 30% sucrose solution in PBS for at least 48–72 hr (until tissue sinks). The 30% sucrose solution was replaced with fresh one every 24 hr. The tissues were completely embedded in Tissue Tek compound (Sakura Finetek, Torrance, CA, USA) for 20 minutes, and freeze at −20°C to −80°C. 10 µm thick tissue sections were carefully cut and collected using a Leica cryostat (Leica Biosystems, Buffalo Grove, IL, USA). The cryostat temperature was between −18°C and −25°C. The tissue sections were mounted on gelatin coated slides (Sigma Aldrich, St. Louis, MO, USA), dried for 30 minutes at RT, and stored at −20°C to −80°C until further use.
The cryostat sections were rehydrated in PBS for 15 min. For epitope retrieval, the sections were immersed into preheated retrieval solution (pH 9.0, 95°C; DAKO, Carpinteria, CA, USA) for 30 minutes and then cooled to RT. The sections were then incubated in blocking buffer (1% bovine serum albumin [BSA] in PBS; Thermo Scientific, Rockford, IL, USA) for 30 minutes at RT to prevent non-specific binding issue on the target epitopes between primary antibodies and tissues. The sections were then incubated with primary antibodies diluted in the diluent buffer (DAKO) overnight at 4°C. Thereafter, the sections were rinsed 3 times for 20 minutes in PBS and incubated with secondary antibodies diluted in the diluent buffer for 90 minutes at RT. For double immunostaining, primary and secondary antibody incubation steps were repeated without the second block step. After this period, the sections were rinsed 3 times for 20 minutes in PBS and then placed in anti-fade mounting media containing 4′,6-diamidino-2-phenylindole (DAPI; Vecta Labs, Burlingame, CA, USA) for 5 minutes at RT to counterstain the nuclei. For staining on negative controls, sections were incubated with the antibody diluent and a non-immune serum (Thermo Scientific) of the same isotype and at the same concentration as the primary antibodies. Confocal microscope (LSM800; ZEISS, Oberkochen, Germany) was used to acquire the images that analyzed by Zen Blue (ZEISS, Oberkochen, Germany).
Epididymis tissues were double-immunostained with ZO-1 and ZO-2, ZO-3, occludin, or Cldn3. A LSM800 confocal microscope was used to acquire images. All images used for quantification were obtained in the same conditions. For comparisons of levels of TJ proteins between groups, the epididymides of three immature and mature animals was conducted and three cryostat sections from each group was labeled and analyzed. The quantification of the expression level of ZO-1 colocalization with each target protein in TJs was calculated by using the Image J software (NIH, Bethesda, MD, USA), as described earlier [31].
The epididymides were homogenized on ice for 30 minutes in RIPA buffer (Thermo Scientific) additive with complete protease inhibitors (Roche, Penzberg, Germany). The homogenized sample was centrifuged at 18,000×g for 20 minutes at 4°C, and then the supernatant were collected. Protein concentration was determined by the bicinchoninic acid (BCA) Protein Assay Kit (Thermo Scientific). Protein samples were diluted with 4x lithium dodecyl sulfate (LDS) Sample Buffer (Bio-Rad Laboratories, Hercules, CA, USA) in the presence of 4% b-mercaptoethanol. Samples were heated at 99°C for 10 minutes and cooled on ice before loading on gels. Proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel electrophoresis and then wet-blotted onto polyvinylidene difluoride (PVDF) membranes for 90 minutes at 90 V. Blots were blocked for 60 minutes with 5% non-fat dry milk in Tris-buffered saline with 0.1% Tween-20 (TBST) at RT. Primary antibodies diluted in TBS were applied overnight at 4°C. After washing in TBST, secondary antibodies were applied for 60 minutes at RT. Antibody binding was visualized by Chemiluminescence reagent (Perkin Elmer Life Sciences) and were detected with iBright FL1500 (Invitrogen).
RESULTS
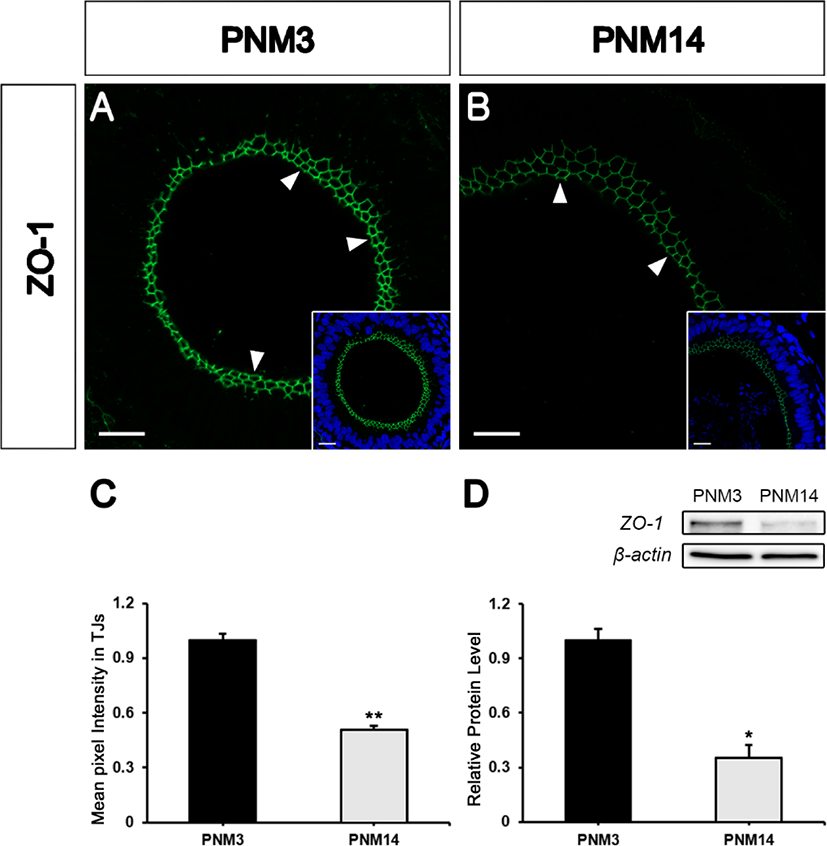
To confirm that ZO-1 is expressed in the goat epididymis, the epididymides at postnatal month (PNM) 3, and 14 were stained with a ZO-1 monoclonal antibody. ZO-1 was strictly expressed in the apical region (where TJs are present) of the epididymal tubules (Figs. 1A and 1B, white arrowheads). The expression level of ZO-1 was higher in the PNM3 than in the PNM14 epididymis. Quantification of the average pixel intensity of ZO-1 immunostaining in TJs showed a drastic decrease in the adult epididymis (Fig. 1C). The same results were observed by western blot analysis (Fig. 1D).
Next, we investigated whether the other two ZO proteins, ZO-2 and ZO-3, were present in the TJs of the goat epididymis. Double labeling with ZO-1 and ZO-2 showed that ZO-2 was co-localized with ZO-1 in TJs of the epididymal tubules (Figs. 2A and 2B, yellow arrowheads). ZO-2 expression pattern was similar to that of ZO-1 (Figs. 2C and 2D, white arrowheads). ZO-2 labeling was brighter in the PNM3 epididymides compared to that at PNM14. Quantification of the average pixel intensity of ZO-2-associated immunostaining in TJs revealed a drastic decrease in the mature epididymis (Fig. 2E).
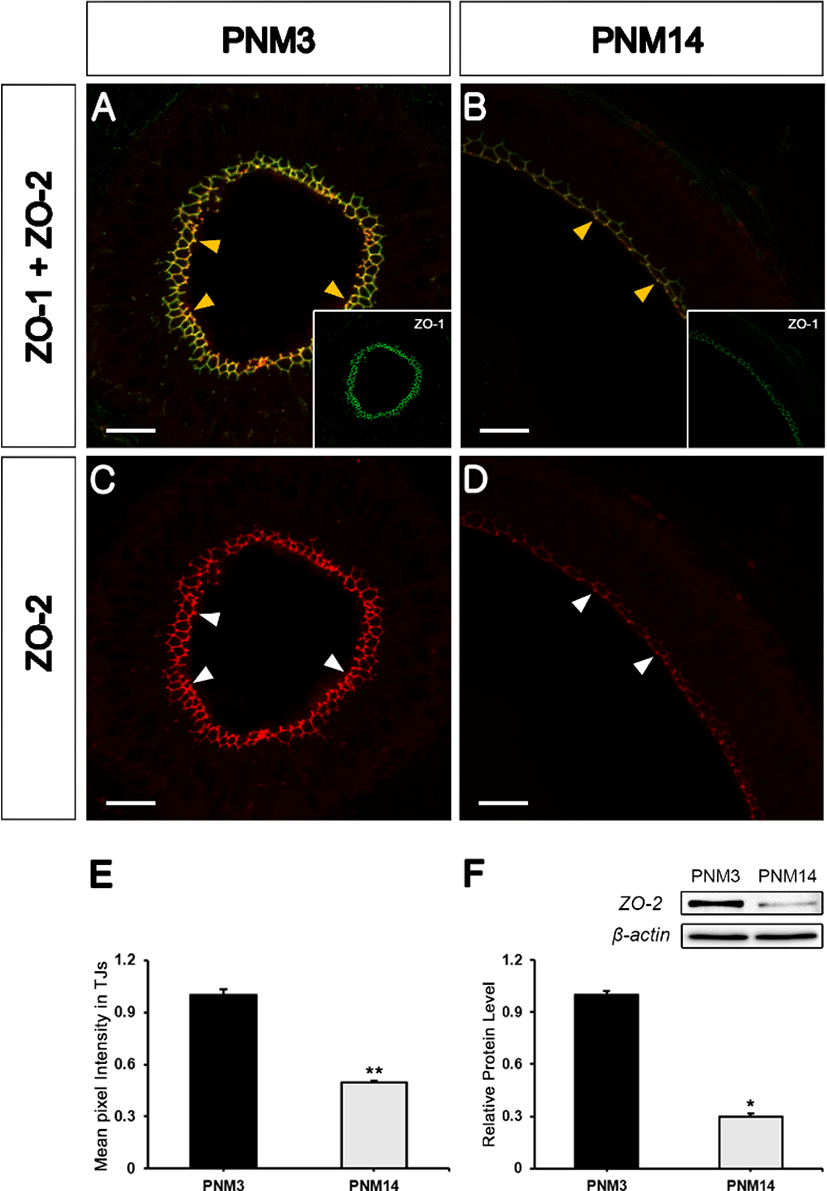
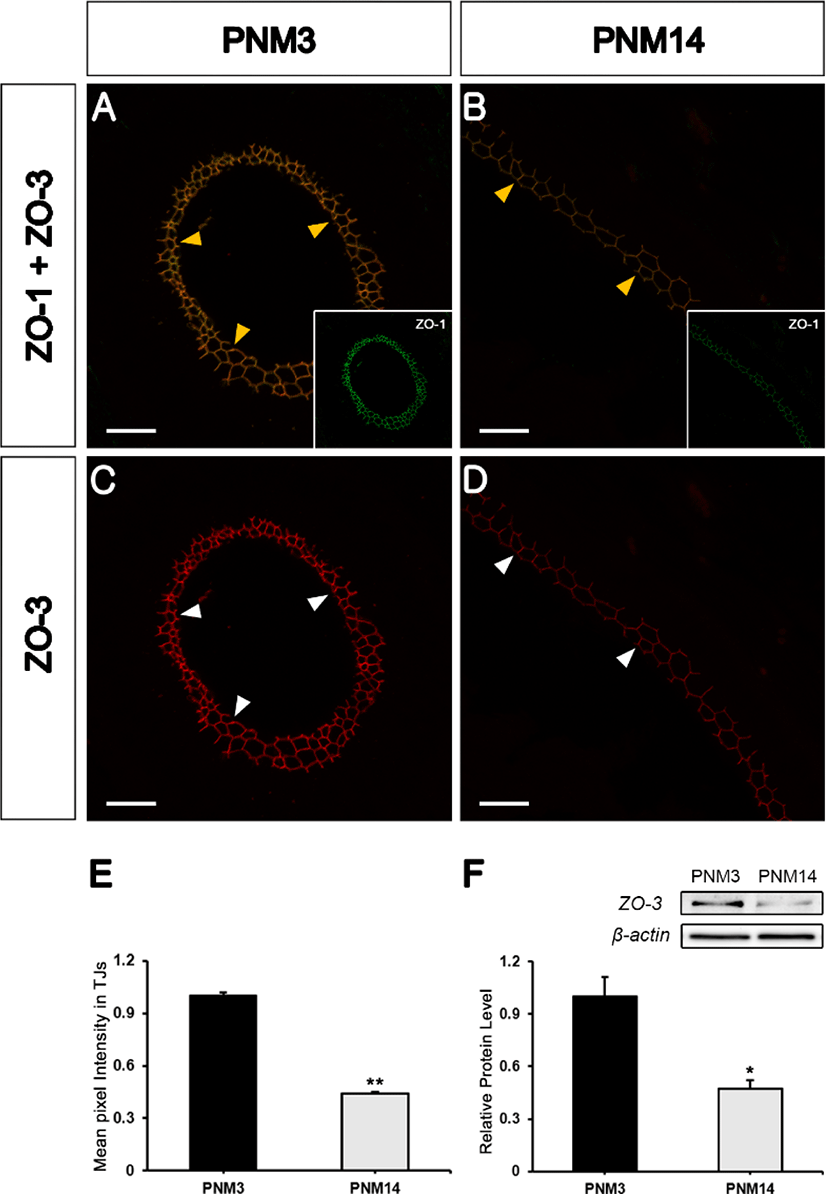
Western blotting verified the decrease in ZO-2 in PNM14 epididymides (Fig. 2F). ZO-3 also colabeled with ZO-1 in TJs in the goat epididymis (Figs. 3A and 3B, yellow arrowheads). Further, its expression was significantly decreased at PNM14 compared to that in PNM3 epididymides (Figs. 3C and 3D, white arrowheads). The quantification of ZO-3 expression was confirmed by immunofluorescence (Fig. 3E) and western blotting (Fig. 3F). These results indicated that all three ZO proteins were maintained at high expression levels during immature stages, but were drastically reduced in the mature epididymides.
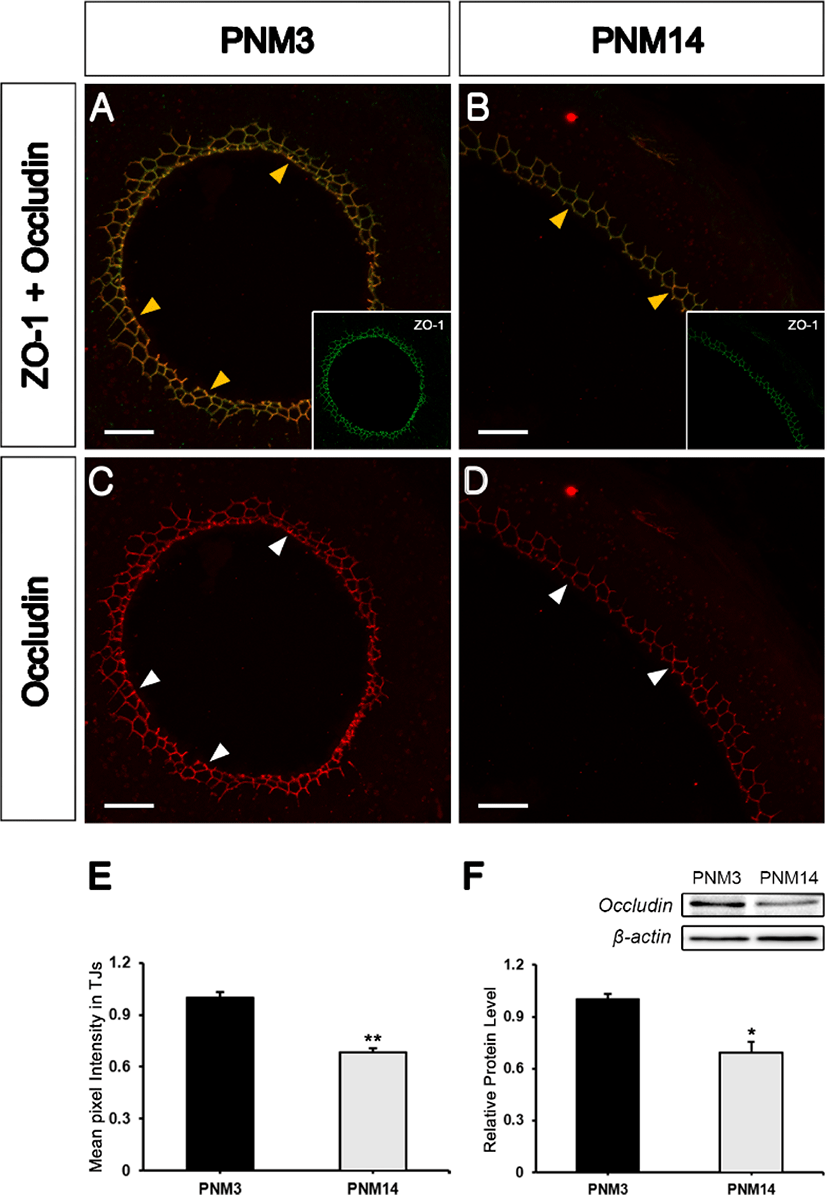
To determine whether occludin, a transmembrane protein, could be localized to the TJs of the goat epididymis, double-staining was performed with ZO-1 and occludin. Occludin protein colocalized with ZO-1 in TJs (Figs. 4A and 4B, yellow arrowheads). Occludin was also significantly decreased in the PNM14 epididymides compared to that of PNM3 (Figs. 4C and 4D, white arrowheads). The quantification of occludin expression was confirmed by immunofluorescence (Fig. 4E) and western blotting (Fig. 4F). This result indicated that occludin was also significantly decreased in the mature epididymis, similar to the three ZO proteins, known as peripheral membrane proteins.
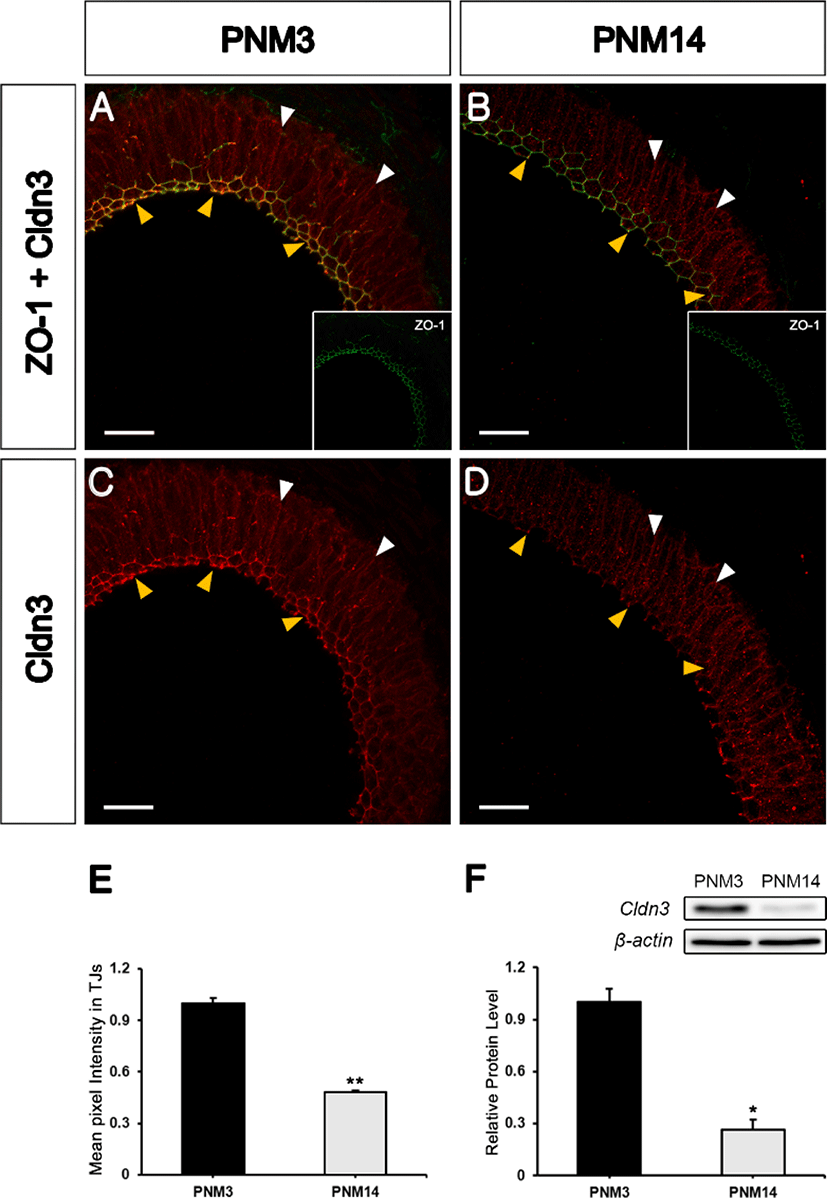
To determine whether Cldn3, a transmembrane protein, could be expressed in the TJs of the goat epididymis, double-labeling was performed using ZO-1 and Cldn3 antibodies. Interestingly, the localization and expression of Cldn3 tended to be age-dependent. At PNM3, Cldn3 was expressed and localized to the TJs (Figs. 5A and 5C; yellow arrowheads) and basolateral membranes (Figs. 5A and 5C; white arrowheads). At PNM14, Cldn3 was significantly decreased in TJs where ZO-1 was localized (Figs. 5B and 5D; yellow arrowheads) and also showed low expression levels in the basolateral membranes (Figs. 5B and 5D; white arrowheads). Quantification of the average pixel intensity of Cldn3-associated immunostaining from TJs revealed a significantly reduced signal at PNM14 compared to that at PNM3 (Fig. 5E). The total expression level of Cldn3 was examined by western blotting (Fig. 5F). These results indicate that Cldn3 is expressed at and localized to the basolateral membrane and TJs. Moreover, the changes in Cldn3 expression patterns appear to be age-dependent.
DISCUSSION
Incomplete spermatozoa can obtain abilities such as progressive motility and fertilization capacity through the final maturation step in the epididymis. The maturation events of spermatozoa in the epididymis depend on a unique luminal environment that is provided and maintained by the formation of the BEB, which is formed by TJs composed of several proteins, including transmembrane and peripheral proteins. These specialized TJ proteins function in forming the seal between epithelial cells, which is essential for sperm maturation as well as sperm protection from the immune system. However, there is still limited information on the regulation mechanism of TJ proteins that are key factors for BEB formation. Here, we determined the expression and localization patterns of TJ proteins, including peripheral membrane proteins, three ZOs (ZO-1, ZO-2, and ZO-3), and the transmembrane proteins, occludin and Cldn3, in the immature and mature goat epididymis.
ZO proteins comprise a family of TJ-associated proteins that have a functional role in linking transmembrane proteins to the actin-based cytoskeleton [21]. In the present study, all three ZOs (ZO-1, ZO-2, and ZO-3) were present and localized along the apical regions of adjacent epithelial cells of both the immature and mature epididymis. These results agree with those of a previous study showing that all three ZOs were present in the TJs of the adult epididymis of mice [31]. In addition, we suggest that in the goat epididymis, their expression levels are different between the immature and mature stages. For example, all three ZO proteins were detected at significantly lower levels in the mature epididymides than in the immature ones (Figs. 1, 2, and 3). Previous studies have demonstrated that ZO-1/ZO-2 and ZO-1/ZO-3 complexes are present, whereas ZO-2 is not associated with ZO-3 [21]. Moreover, we previously demonstrated the protein expression compensatory relationship between ZO-2 and ZO-3 without affecting ZO-1 expression levels in the epididymis of mice [31]. However, in the present study, we did not observe any compensation between ZO-2 and ZO-3 during goat epididymis development. These unexpected findings suggest the possibilities that other TJ proteins could contribute to BEB formation in the mature epididymis instead of ZO proteins or that specific relationships among these three ZO proteins might create an optimal system in this organ. Further studies are needed to clarify the reason underlying the reduction of all three ZO proteins in the mature epididymis.
We also observed that occludin and Cldn3 proteins, transmembrane proteins associated with ZO-1, ZO-2, and ZO-3 were localized and expressed in TJs. The localization and expression of a few types of transmembrane proteins in the epididymis have been previously reported in various species, including pigs [32], canines [33], rabbits [34], mice [31], and rats [35]. Occludin expression in TJs appears from embryonic day 18.5 in the mouse epididymis and its expression is maintained until adulthood [36]. Various Cldns were also observed in the epididymis, including Cldn3, which is expressed in the TJs and basolateral membranes in mice [31], rats [37], and bats [38]. In agreement with these previous studies, we found that occludin was present in TJs and that Cldn3 was expressed in both the basolateral membranes and TJs in the goat epididymis. Moreover, our results indicated much higher expression of occludin and Cldn3 in the immature epididymis, as high levels of ZO proteins were also observed. This might be because the high expression of ZO proteins in the immature epididymis can increase the chance of an association with occludin and Cldn3. Otherwise, the reduction of ZO proteins could result in a decreased chance of binding to other transmembrane TJ proteins, such as occludin and Cldn3. All TJ proteins examined in this study were expressed at significantly higher levels in the immature epididymis than in the mature epididymis. These findings suggest that some factors regulating the expression of TJ proteins are likely to be present during epididymis development. Additional studies investigating the expression and localization patterns of other TJ proteins are needed to elucidate these unexpected issues in the goat epididymis.
Testicular luminal factors (TLFs) might be important in regulating changes in the distribution of TJ proteins. TLFs synthesized and secreted from the testis, include hormones, growth factors, and other luminal nutrients, which are directly in contact with epididymal epithelial cells that contribute to the formation of TJs and play an important role in the development of the epididymis [1, 39-41]. According to a previous study, the secretion of testicular luminal fluid might occur only in the normal mature testes of many species [42]. Therefore, it is possible that the epithelial cells exposed to TLFs during puberty might contribute to changes in the levels of TJ proteins in the mature epididymis. We previously reported that the epithelium is not perfectly differentiated until PNM2 [43] and PNM3 (unpublished data). Thus, the undifferentiated epithelium has a stronger TJ association between epithelial cells, and those that are differentiated by contact with TLFs could reduce their TJ association at the epididymal epithelium in goats. The reduction of these TJ proteins might modify the epididymal luminal contents that make up a unique environment in the adult epididymis for sperm maturation and storage. In addition, among TLFs, an androgen hormone known to be the most important key factor is responsible for regulating and maintaining epididymal structure and function. Although not well understood, an association exists between TJs and androgens. Several studies have shown that androgens regulate TJ molecules including Cldn3, Cldn11, occludin, and ZO-1 [44-46] and are crucial for the regulation of every aspect of spermatogenesis via formation of the BTB [47,48]. However, direct links between androgens and TJ proteins in the epididymis have not been well documented. Therefore, more research on the effect of androgens on BEB formation is needed to confirm this assumption. Moreover, estrogen, another steroid hormone, is a good candidate factor that controls the expression and localization of TJ proteins. Estrogens are known as the most important female hormone, but in males, estrogens are critical to maintain testicular and epididymal function [49,50]. Without estrogens, male animals are infertile owing to failed fluid reabsorption, which is necessary for sperm concentration maintenance in the efferent ductules of the epididymis [51]. Earlier studies also provided some evidence that estrogens negatively control BTB function. Exposure to estrogens or diethylstilbestrol (a synthetic nonsteroidal estrogen) delays BTB formation and induces spermatogenesis failure in rats [27,52]. Moreover, bisphenol A (an estrogenic environmental toxicant) treatment induces disruption of the TJ barrier by changing the expression and localization of transmembrane proteins at the BTB [53]. Therefore, it is possible that an increased exposure to estrogens in the adult epididymis might decrease the expression levels of TJ proteins in the BEB.
Based on the present results, ZO-1, ZO-2, ZO-3, occludin, and Cldns are expressed and localized at the TJs of immature and mature epididymides of goats. Moreover, all TJ proteins were expressed at much higher levels in the immature epididymis than in the mature epididymis. This finding implies that undifferentiated epithelial cells might form a stronger BEB through high expression levels of TJ proteins, and then, the expression of TJ proteins decreases with the differentiation of epithelial cells in the goat epididymis. Our findings could contribute to a better understanding of the organization of TJ proteins in the BEB during the development of the goat epididymis. Although we could not directly explain the regulatory mechanism of TJ proteins in the BEB, our findings further the understanding of the reproductive physiological characteristics of the goat epididymis.