













INTRODUCTION
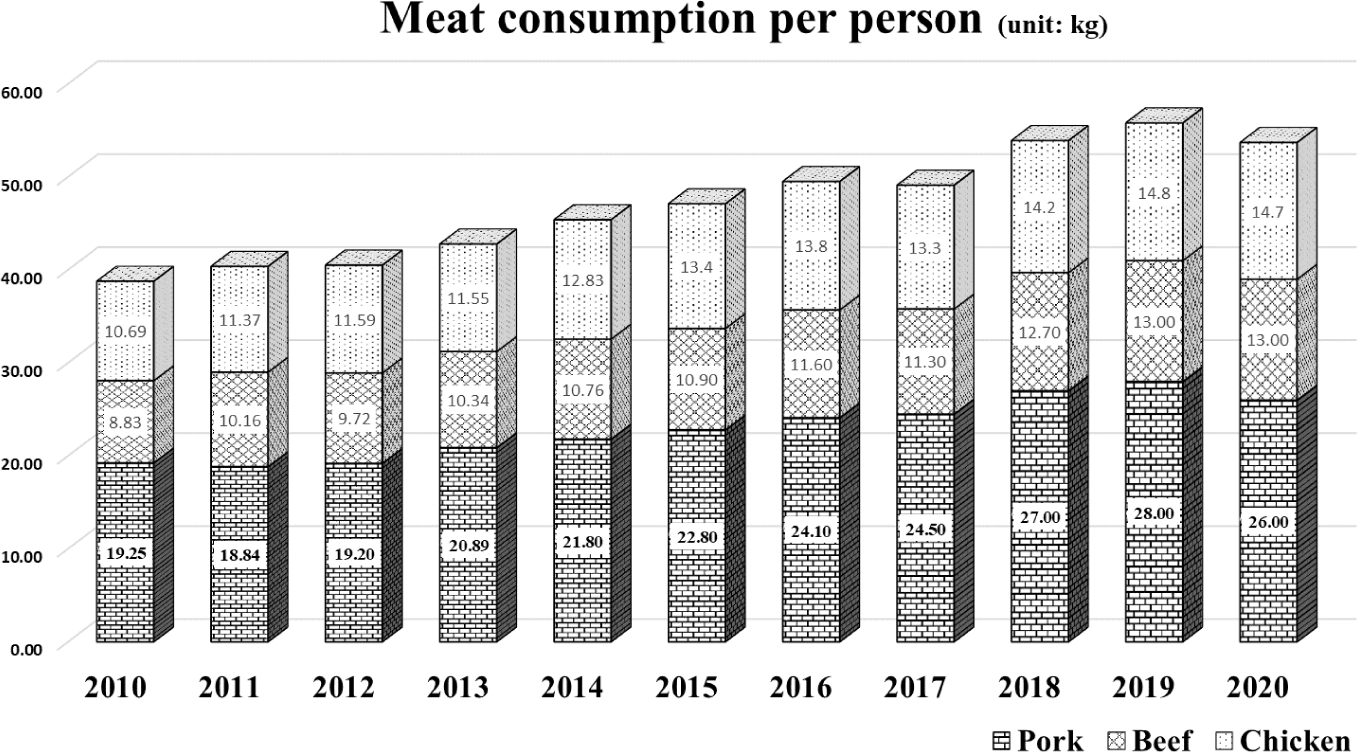
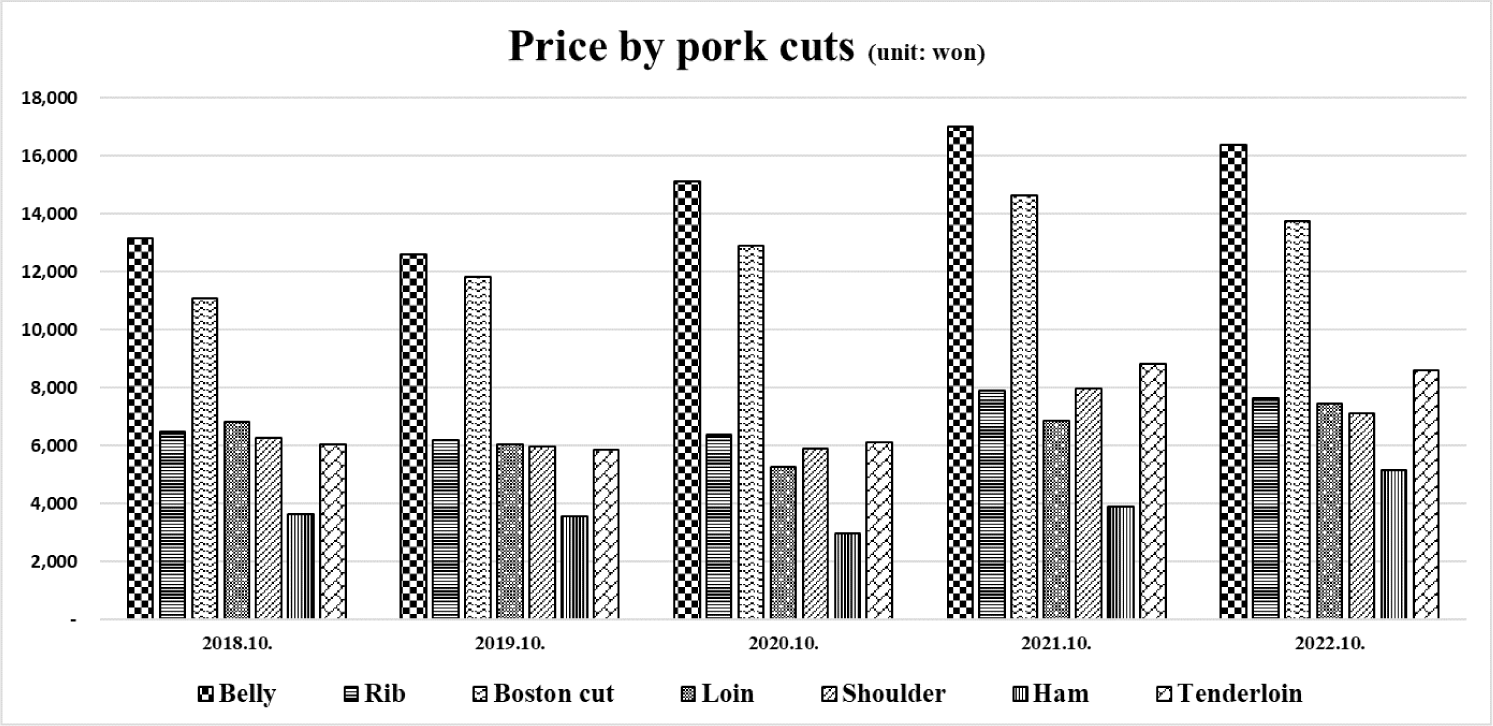
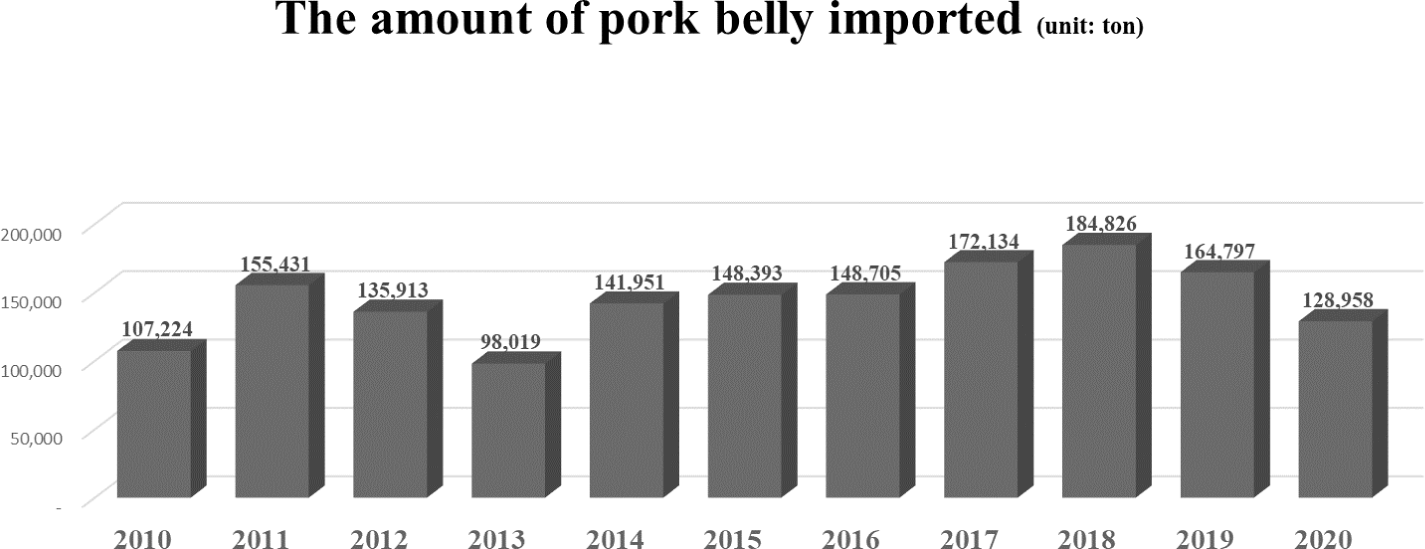
Pork, one of the most consumed meats, has been preferred by consumers [1]. It has been particularly recognized as an essential source of animal protein [2]. According to that preference, meat consumption per person has been steadily increasing (Fig. 1). Among the many parts of meat in pork, the belly, known as high-fat cutting and, has been preferred by consumers in South Korea [3–5]. The price of the belly is the highest than the other pork meat (Fig. 2). This means that the belly is the most preferred cut of pork in South Korea. Pork belly is imported in South Korea because it cannot meet the demand with domestic production, and the import volume is increasing tendency (Fig. 3). Most belly is consumed via a roasting cooking in South Korea called ‘Samgyeopsal’ in Korean word. Moreover, in East Asia, pork belly is also a preferred part of meat as various cooking ingredient compared to the Western for bacon [6]. Therefore, the belly seems to be a large part of the pork market worldwide, and its marketability is expected to expand compared to the present.
Pork belly, which has copious flavor and taste, has known as a high-fat cut among primal pork cuts because of its high subcutaneous fat [4]. Nevertheless, too much subcutaneous fat composition derives a greasy taste, which makes it a non-preferable factor [5,7]. The high-fat belly (also known as ‘caky-fatty’), a non-preferred belly cut, appeared in the 5th lumbar vertebra and the 12th thoracic vertebra with high subcutaneous fat [7]. However, Koreans prefer the fat cut more than Europeans [3]. According to those reports, that is a fact pork belly is the favored meat in South Korea and other countries. Moreover, its marketability is a large scale to focus on by the swine industry, including South Korea. However, the standard belly cut of the consumer preference was different. In addition, the pork evaluation system in South Korea has focused on lean meat production, represented by loin muscle traits and back fat thickness, except for the belly. In this study, we reviewed the previous reports to focus on the characteristics of pork belly for improving heading to consumer preference in many ways, including phenotypic and genetic approaches and the possibility of improving using animal breeding in South Korea.
THE CHARACTERISTICS OF PORK BELLY
Pork belly is officially defined that “The abdominal muscle from the fifth rib or sixth rib to the last lumbar spine (including the navel and dorsal oblique muscles) after separation from the fat mass of the humerus at the hind leg along the thin membrane of the torso and abdominal muscles” in South Korea (Fig. 4) [11]. The pork belly contains some component muscle and fat, comprising 55%–60% adipose tissue [12]. The belly fat is divided into two layers: subcutaneous and intermuscular fat. The significant component muscles of the belly are designated as the cutaneous trunci muscle, latissimus dorsi muscle, pectorales profundi muscle, rectus abdominis muscle, external abdominal oblique muscle, serratus ventralis, diaphragm, intercostal externi, and obliquus abdominis interni, and the others.
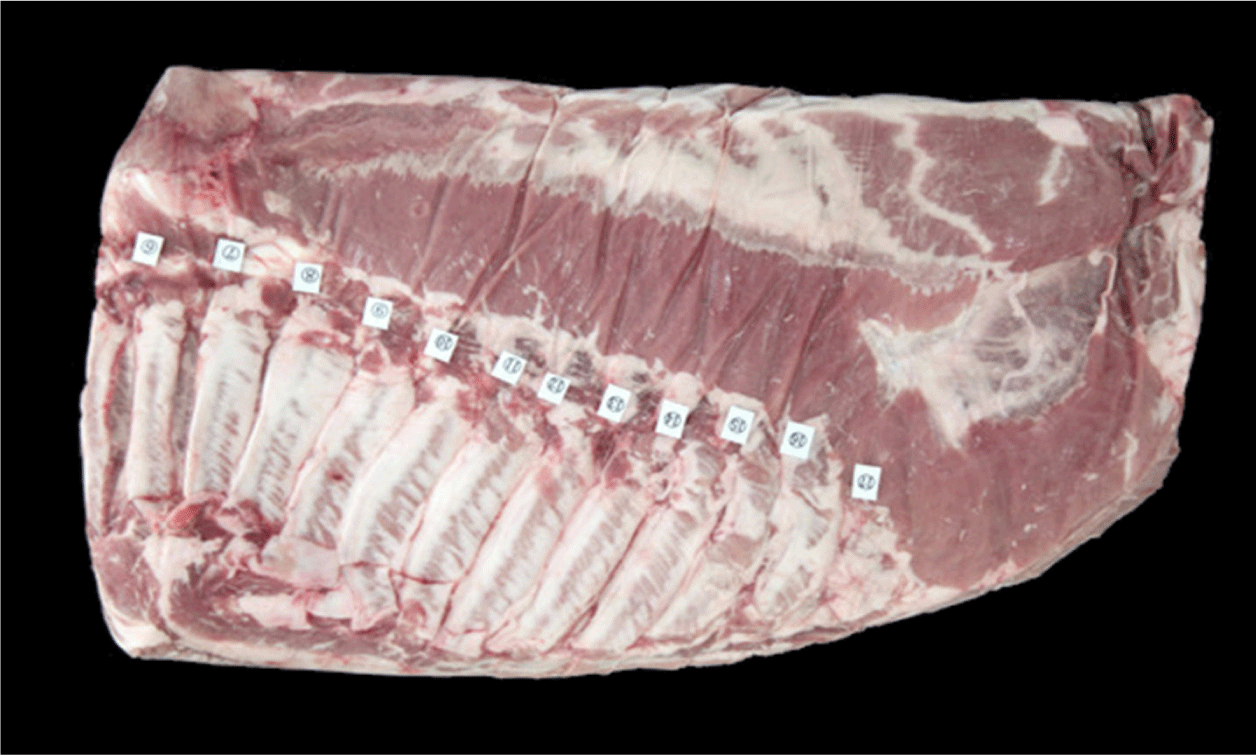
Kim reported that the pectorales profundi muscle showed a characteristic of a constant decrease in the thoracic vertebrae area of the belly and then disappearing within them [7]. The cutaneous trunci muscle, a significant component muscle of the belly, steadily increases from the thoracic vertebrae and is observed in the lumbar vertebrae area of the belly. The latissimus dorsi muscle, developed above the cutaneous trunci muscle, was majorly observed at the beginning of the thoracic vertebrae area of the belly. The rectus abdominis muscle is irregularly developed from the 5th–6th thoracic vertebrae, and its maximum area is majorly watched at the end of the thoracic vertebrae point of the belly. The external abdominal oblique muscle is also developed, like the rectus abdominis muscle, to the end of the thoracic vertebrae. It rapidly disappears at the beginning of the lumbar vertebrae area of the belly.
The measurement method of the belly was previously introduced to a passive and an automatic methods. A passive method means directly measuring using a ruler and a scale. A grader of the Animal Products Grading Service (APGS) used that method for a long time [13]. However, as the slaughter amount of pork has increased, that method has become less efficient because of the required time for measurement [14]. The automatic measurement system (AMS) was developed because of these problems.Automatic Fat-O-Meat’er (AutoFOM), one of the AMS has been distributed in Europe since the 1990s, and after that, VCS2000, the visual analyzing system, was developed and distributed [14]. The information on the instrument is summarized in Table 1.
| Instrument | Producer | Measuring principle | Reference |
|---|---|---|---|
| UltraFOM (UFOM, UltraFOM 300) | SFK Technology | Ultrasound | 15–17 |
| AutoFOM | SFK Technology | Ultrasound | 15–17 |
| CVT-2 | AUS | Ultrasound | 15 |
| Vision-Based Video Image Analyzer (VCS2000) | E+V Technology GmbH | Video image | 12,13,17 |
The FOM method classified automatic and manual systems [15]. The FOM measurement system measured lean meat percentage (LMP) and fat thickness of the carcass including belly region using ultrasonic instrument [16,17]. The AutoFOM system, based on the ultrasonic scanner, has 16 ultrasonic sensors, and it measures the carcass at an interval of 5 mm 200 times during the slaughter process [18]. The AutoFom reported that it showed a lower ratio of error than the other non-destructive automated inspection in the European standard [16]. UltraFOM, another FOM method system, handled manually as non-invasive ultrasound instrument unlike AutoFOM [15]. The FOM method is easy to measure the pork belly parameters, because of ultrasonic.
VCS2000 is an instrument which automatically measured LMP from the half of carcass including belly region via video based image measurement [18,19]. Font et al. previously reported that the estimation accuracy of LMP via VCS2000 was lower than FOM and AutoFOM system [16]. However, an effort to decrease accuracy differences between FOM method and VCS2000 was proceeded via correcting estimation equation [13]. Moreover, the differences were derived from the variation of evaluation system, hence it is necessary to identifying an equation which fitted an environment of evaluation.
CONSUMER PREFERENCE FOR THE PORK BELLY
People consumed pork belly as bacon for roasting in the Western world. However, South Korea’s pork belly is consumed as a raw meat shape for roasting or steaming. In the early, the muscle composition occupied 22%–23% of the pork belly [20]. Consumers demand increasing meat composition for bacon [20]. Stiffler et al. [21] also reported that the muscle-fat ratio could affect consumer preference. According to this preference, the pork belly fat region decreases by around half [6,12]. However, it affects fat separation and other sensory issues [22]. The increased belly-muscle ratio has been derived from increasing the moisture content and polyunsaturated fatty acid (PUFA) [23,24]. As a result of the swine breeding to increasing muscle ratio, the product of the pork belly becomes soft and thin [24]. Hence, it has been challenging to handle the processors. Moreover, the soft and low-fat bellies may cause to reduce storage period and poor product quality [6].
There were three sights against the pork belly: producers, processors, and consumers. From the producer’s point of view, the quality of the belly is the same as its weight [6,25]. It is because why pork meat is priced by its weight. Furthermore, the viewpoint of meat processors prefers heavy belly weight and thick belly for processing [23]. Moreover, a thick belly is known to relate to firmness because of belly fat composition, including a low PUFA ratio [24]. However, the increased pork belly weight may cause a concomitant rise in fat composition. Therefore, it is important to find a suitable muscle-fat ratio. From the point of view of consumers, the nutritional and appearance parameters such as flavor, total fat, fatty acid composition, color, and thickness [12]. Person et al. additionally reported that consumers prefer to thin and average belly thickness as bacon slices [12]. However, since it is a perspective of consuming bacon, this preference may be different from that of Korea, which consumes grilled belly.
In South Korea, fat composition (53.4%), meat color (25.8%), wideness (14.5%), and fat color (4%) of the belly parameters were attractive traits by consumers [7]. In addition, another study reported as the numericalized data that the consumer preference of the belly for thickness was approximately 3.94 cm [26]. Vonada et al. also presented other belly parameters, including fat contents, lean color, and belly weight, which Korean consumers preferred. Korean consumers preferred a moderate amount of moderate fat contents (approximately 20%–40%) and 4.04 kg of the belly weight [26]. As it is the most consumed area in Korea, many preference surveys were expected to be conducted, but few showed the exact numericalized data. In addition, since the focus was only on meat color (lightness), fat ratio, and belly weight, it was difficult to investigate the detailed muscles that makeup pork belly and the characteristics of each muscle. However, based on previous studies, improvement should be carried out with the goal of breeding pork belly.
THE DETERMINING FACTORS FOR THE BELLY IN THE GROWTH STAGE
The growth stage for the pork belly, which is made up of various muscles and fat, can be divided into three parts: myogenesis, fat deposition, and nutritional background. At first, myogenesis is well reported by previous studies. The myogenesis started at the embryo stage to post-natal [27–29]. During the embryonic development stage, mesoderm started myogenesis with the first muscle fiber construction, and the muscle fiber proliferated to the additional fibers [29,30]. The proliferation of the myogenic progenitors become active in the initial stage, whereas the activity decreases as the reaches a steady state of the number of myonuclei [31,32]. From the viewpoint of pork belly, lateral trunk and limb muscles are associated with pork belly muscle characteristics. The lateral trunk and limb muscles were reported to be derived from the hypaxial domains during embryo development [30].
Fat deposition is derived from adipogenesis. The adipocyte is divided into two central depots: subcutaneous adipose tissue and visceral adipose tissue in the human study [33]. These adipose tissues are similarly observed and measured in the livestock animals such as swine and cattle [34]. Pig adipose tissue developed during the cellular hyperplasia stage between 7 and 20 kg [35]. Moreover, some studies reported that the intermuscular fat, composed of pork belly, showed different growth patterns against their anatomical location [36,37].The growth rate of intermuscular fat in the belly is more rapid than subcutaneous fat, whereas the ham observed reverse growth [36,37].Another previous study backed up these observations that 18% of intermuscular adipose tissue develops in the pig growth stage due to 23 kg of body weight (from weaning to post weaning) [38]. It then decreases its ratio to 13% due to 114 kg body weight. The fat ratio presented 66% at the 91 kg body weight based on 100% at the 114 kg [38]. Therefore, the identification of intermuscular fat regulation factors affecting the rate of development against subcutaneous fat in pre-finishing pigs is needed.
The nutrition factor is another important point of pig growth and its belly site. Short chain fatty acid (SCFA), a product of bacteria fermentation, was reported that plays a role in providing energy for host cells as gut microbiota [39]. In the pig study, oral administration of SCFA could be affected to decrease fat deposition [40,41]. Another fat deposition study reported that the ratio of Archaeal species with methanogenesis abundance, deriving high-fat deposition, in high-fat pigs was higher than in low-fat pigs [42]. Moreover, butyrate-producing bacteria species, improving SCFAs, was identified in the low-fat pigs [42]. High fat diet could associate with abdominal fatty acid deposition in abdominal fat in pigs [43]. In addition, Duroc pigs had a resistant to fatty acid composition of the diet [43]. Therefore, low-fat diet could be affected to decrease abdominal fat deposition, whereas further study with pig microbiome which relate to fat deposition microbiota is needed to identify decreasing belly fat deposition to maintain firmness of pork belly.
For a long time, an effort for improving production ability has been continuously conducted as a goal of pig breeding. The production trait was traditionally classified as loin eye area, as a representative skeletal muscle, growth performance, and back fat thickness. In addition, a carcass weight and live weight is known to use to estimate carcass composition and its related muscle and fat composition [44]. The skeletal muscle has been known to one of the major factor of the carcass [45]. Especially, muscle fiber characteristics consist of the skeletal muscle such as total number of fiber, size of muscle fiber were reported that related to lean meat production ability [45–48]. Moreover, the dimensional pork parameters including size, thickness, and weight were recognized by meat processors as the pork belly production ability [23]. However, the lack of study for pork belly component muscle as a lean meat production ability. Pork belly consist various muscles, which are composed of muscle fiber characteristics. Therefore, it is necessary to identify its characteristics in the pork bellies.
Some studies reported the phenotypic correlations between pork belly components and lean meat production traits. Hermesch reported that the fat percentage of the belly had a negative correlation with the rib bone-muscle area (−0.34) and had a positive correlation with the intermuscular and subcutaneous fat area (0.63 and 0.66, respectively) [49]. Moreover, the other phenotypic correlations between the rib bone-muscle area and other belly traits were slightly positive. Miar et al. also performed the phenotypic correlation between carcass and meat quality traits in commercial crossbred pigs [50]. The relationships with untrimmed belly weight against hot carcass weight, back fat depth, loin depth and loin eye area were estimated (0.58, 0.31, 0.12 and 0.39, respectively). Another phenotypic correlation between carcass traits and pork belly components has been reported: the seventh slice of the belly components positively correlated with whole belly traits [51]. However, these correlation studies did not estimate using all component traits of the belly. Therefore, further study is needed to estimate the phenotypic correlation among parameters, including pork belly components.
Meat quality traits was roughly categorized to sensory quality (i.e. visual texture and flavor), technological quality (i.e. water holding capacity and pH), and nutritional quality (i.e. protein contents, vitamins and minerals) [52]. In addition, the sensory qualities such as tenderness, juiciness and flavor was importantly recognized by the consumers [53]. Intramuscular fat level was also positively recognized as a factor for negotiation of eating quality (i.e. juiciness, tenderness, flavor intensity and oily mouth feel) [54,55]. As the biochemical constitution of muscle, muscle fiber characteristics reported to affecting meat qualities such as meat color and pH [56–58]. In the pork belly, belly firmness has been known to the quality parameter for processors [23]. Therefore, sustaining the meat quality and improving pork belly quality is important for consumers.
Some researchers have reported the phenotypic correlation between belly parameters and meat quality. A previous study reported that the estimated correlation between belly yield and meat quality did not show significance [59]. Another study presented that the phenotypic correlation between pork belly and meat quality showed weak correlation coefficients [49]. Miar et al. estimated the phenotypic correlation between belly weight and meat quality [50]. The trimmed belly weight obtained a weakly negative correlation with cooking loss, shear force and pH (−0.08, −0.12 and 0.08, respectively). In addition, untrimmed belly weight significantly correlated with loin muscle lightness (0.13). The other meat quality traits did not show significance. The phenotypic correlation among the belly weight, yield and meat quality traits had a weak relationship. Moreover, other belly traits, including component parameters, did not estimate. It is necessary to estimate between belly components and meat quality traits.
THE GENETIC POTENTIAL FOR IMPROVING PORK BELLY TRAITS
Estimating genetic parameters are needed to use pork belly parameters to improve swine breeding. In addition, since the impact on the industry may vary depending on the difference in breeding goals, it is necessary to set the correct target traits. Do reported that part meats’ weight, such as Boston cut and bellies associated with plant age [60]. Moreover, it was also suggested that which traits selected for the goal of swine breeding could affect the pork industry. If the goal were focused on the weight of the belly, the pig would be changed bigger, whereas focusing on the meat cut percentage will change the body shape of the pigs. Therefore, genetic parameters such as heritability and genetic correlation were estimated by some studies.
Hermesch et al. reported that the heritability of pork belly for lean meat was 0.23 (intermuscular fat area) to 0.34 (fat percentage) [49]. The heritability of the belly weight was presented as 0.27 to 0.31 in another study [61]. Willson et al. reported estimated genetic parameters for pork quality traits, including belly width and weight [62]. Those studies were width, fat area, and total muscle area of the specific parts of the belly. The reported heritabilities appeared moderate. Therefore, improving pork bellies for consumer preference, heading to less fat and increasing muscle ratio, could be possible via swine breeding. However, the reported heritabilities of belly traits did not vary. Therefore, further estimation of heritabilities for detail traits such as the area or volume of component muscle and muscle and fat ratio is needed. Kang et al. [63] reported heritabilities using detailed belly traits; it showed moderate to high (0.27 to 0.49), but its population size was small; thus, a more extensive population study is needed to improve accuracy.
The genetic correlation among the pork belly parameters such as rib bone-muscle area, fat ratio of the belly, intermuscular fat area and subcutaneous fat area has been reported to −0.24 to 0.84 [49]. Do et al. [64] reported that the genetic correlation showed moderate to high correlation coefficient among carcass traits including belly weight (0.88 for carcass weight, 0.46 for back fat thickness and 0.80 for LMP). In the commercial cross bred study, trimmed belly weight genetically correlated with weaning weight, average daily gain, back fat thickness and intramuscular fat [50]. The previous study indicated that fat-associated traits had genetically high relatedness. However, the traits did not divide to detail for instance, pork belly component muscle area hence, it is necessary to the further study among the belly component.
The associated genes for pork belly presented in Table 2. There were some candidate genes reported to associate with pork belly parameters. Moreover, the abdominal site, which is the same as the pork belly region, is recognized as a key for obesity studies in humans. Therefore, obesity-associated genes were reported to cause fat deposition in the swine. With the availability of genetic analysis at the DNA level, RYR1 (also known as halothane gene) has been reported to the relationship with carcass composition traits, including fat tissue development in pigs [65,66]. Fat mass and obesity-associated (FTO) gene, a representative obesity-associated gene in the human study, reported that related to pork abdominal fatness in Meishan × Pietrain F2 pigs, including abdominal fat weight (AFW) and backfat thickness, whereas its average daily gain did not significant among the genotypes [67]. Other studies using other breeds supported that the FTO affected AFW [68–70]. High mobility group AT-hook 1 (HMGA1) and melanocortin 4 receptor (MC4R) were reported to associate with a fat deposition measurement in pigs [71,72]. However, other studies reported that MC4R was insignificant [73,74]. The genome-wide association studies (GWAS) result previously presented that ELOVL6, SCD, and FASN affected fat deposition traits [75]. miR-130a reported suppressing Peroxisome proliferator-activated receptor gamma (PPARG) gene expression; hence, the preadipocytes were inhibited. The inhibited preadipocytes made a difference in fat deposition between intramuscular and subcutaneous fat [76]. cAMP-responsive element-binding protein (CREB)-regulated transcriptional coactivator 3 (CRTC3), well known to be related to obesity in humans, has been reported to associate with intermuscular fat thickness, total muscle area and total fat ratio in pork belly [77,78]. However, the study of genes with component muscles in pork belly did not report yet. Therefore, it is necessary to use a genetic approach, including GWAS and Omics analysis for pork belly component muscles.
| Gene | Trait | Species | Reference |
|---|---|---|---|
| ADCY8 | Abdominal visceral fat | Human | 85 |
| ALDH | Fat deposition, abdominal fat contents | Human | 26,82 |
| ATXN1 | Adipocyte development | Human | 83,84 |
| BBS9 | Abdominal visceral fat | Human | 85 |
| CRTC3 | Intermuscular fat thickness, fat ratio | Pig | 77,78 |
| DNAJC1 | Abdominal subcutaneous adipose tissue | Human | 85 |
| EBF1 | Adipocyte development | Human | 83,84 |
| EBLN1 | Adipocyte development | Human | 83,84 |
| ELOVL6 | Fat deposition | Pig | 75 |
| ENSA | Adipocyte development | Human | 83,84 |
| FASN | Fat deposition | Pig | 75 |
| FTO | Abdominal fat weight | Pig | 67–70 |
| GRAMD3 | Adipocyte development | Human | 83,84 |
| GSDMB | Voumetric subcutaneous fat | Human | 83,84 |
| HMG1A | Abdominal fat contents | Broiler | 71,72,80,82 |
| IRS1 | Abdominal subcutaneous adipose tissue, WHR | Human | 87,88 |
| ITPR2 | Abdominal subcutaneous adipose tissue, WHR | Human | 87,88 |
| KCNK9 | Abdominal visceral fat | Human | 85 |
| MC4R | Fat deposition | Pig | 71–74 |
| miR-130a | Suppressing PPARG | Pig | 76 |
| MLLT10 | Abdominal subcutaneous adipose tissue | Human | 85 |
| PPARG | Fat deposition, abdominal fat contents | Pig, broiler, human, mice | 26,80,82 |
| RREB1 | Adipocyte development | Human | 83,84 |
| RSPO3 | Abdominal subcutaneous adipose tissue, WHR | Human | 87,88 |
| RYR1 | Fat deposition | Pig | 71,73 |
| SCD | Fat deposition | Pig | 75 |
| SPARC | Subcutaneous adipocyte deposition | Cattle | 76 |
| STAB1 | Abdominal subcutaneous adipose tissue, WHR | Human | 87,88 |
| TBX156 | Abdominal subcutaneous adipose tissue, WHR | Human | 87,88 |
| UBE2E2 | Adipocyte development | Human | 83,84 |
| WARS2 | Abdominal subcutaneous adipose tissue, WHR | Human | 87,88 |
| WNT family | Myogenesis, adipogenesis | Broiler | 79 |
The overall genes were presented in Table 2. In other livestock, bellies did not consume part meat. Therefore, the genetic approach against belly meat was not enough. However, in the myogenesis described above, belly muscles were developed by hypaxial domains during embryo development [30]. In addition, adipogenesis, associated with fat deposition, is also a crucial factor for the quality of bellies, and previous studies knew its associated genes well. Wnt gene groups were reported to relate to cell fate and development as the associated factor with myogenesis and adipogenesis [79]. PPARG and HMG1A were reported to relate abdominal fat contents in a broiler study [80]. The beef cattle study reported that SPARC gene in subcutaneous adipose tissues overexpressed to compare with intramuscular adipose tissues [81]. Of the gene expression profiling analysis between subcutaneous and intramuscular fat, 7,526 genes were commonly expressed, whereas only 12 genes were specifically expressed in subcutaneous fat [34]. The differently expressed gene network between subcutaneous and intramuscular fat reported that PPARG and ALDH were observed as key genes [34,82].
In the human study, a previous study reported that ATXN1, UBE2E2, EBF1, RREB1, GSDMB, GRAMD3 and ENSA related to adipocyte development using GWAS meta-analysis [83,84]. Moreover, the GSDMB is related to volumetric subcutaneous fat. Another GWAS presented that BBS9, ADCY8 and KCNK9 were associated with abdominal visceral fat, and MLLT10, DNAJC1 and EBLN1 near SNPs related to abdominal subcutaneous adipose tissue [85]. In a race family-based study, a previous Genome-wide linkage scan presented that several loci, such as 2q22.1 and 2q33.2-q36.3 region (IRS1 locus), obtained significance [86]. The functional studies reported that fat deposition-related genes such as RSPO3, TBX156, ITPR2, WARS2 and STAB1, which are known to be associated with waist-hip ratio differently expressed in abdominal subcutaneous adipose tissue [87,88]. Based on these GWAS results, it is necessary to apply swine breeding for the pork belly component.
CONCLUSION
Pork belly, constructed with many muscles and fat, is a highly consumed part of meat in South Korea as a roasting cooking. In addition, the price is maintained high because of high demand. Most of the swine for breeding in South Korea was imported from the Western world, where the established evaluation standard focused on the loin muscle. Nevertheless, the pig breeding and pork evaluation system in South Korea was only focused on the loin muscle area, the same with the Western world. The belly consists of intermuscular and subcutaneous fat and various muscles. However, the genetic parameters of pork belly have yet to be estimated, such as its component parameters. To use to available in swine breeding, it is necessary that the estimation of genetic parameters and phenotypic correlations against whole belly components. Moreover, a genome-wide approach is required to identify associated genes against belly parameters to use genomic selection. Based on available data in public domain, pork belly, the highest consumption as a grilled of other pork cuts in South Korea, could be available to use for breeding as an economic trait via genomic approaches.