
INTRODUCTION
Feather and skin color distinguish poultry breeds and are important morphological characteristics. The plumage color of breeds is strongly related to economic traits such as growth rate, egg production, and carcass traits [1,2]. For example, white plumage is preferred in broiler chickens because feathers from white chickens are easier to remove than those from colored chickens [3]. The Yeonsan Ogye is a Korean chicken that is known for its black feathers, skin, bones, and comb [4]. This is an uncommon phenotype, and the Yeonsan Ogye has been registered as a natural heritage of Korea (no. 265). Selection is used to maintain the standard phenotype of the Yeonsan Ogye chicken, as individuals with various other plumage colors are occasionally observed [5]. Exceptional individuals are eliminated from the breeding pool to maintain the natural heritage phenotype.
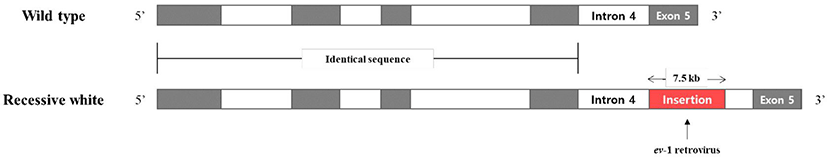
Plumage color in birds is determined by pigments such as carotenoid and melanin [6,7]. Melanin is involved in the expression of black (eumelanin) and red (pheomelanin) and is synthesized by tyrosinase [8]. Two autosomal loci are associated with white plumage color in chickens [9]. The PMEL17 gene gives rise to a dominant white (locus I) and has three alleles (dominant white I, smoky IS, and dun ID) [10]. The tyrosinase (TYR) gene is associated with a recessive white (locus c) and also has three alleles (recessive white c, albino ca, and red-eyed white cre) [11]. The recessive white mutation (c) is reported to involve insertion of the avian leukosis virus ev-1 sequence at the end of intron 4 in the TYR gene [12], which inhibits the transcription of exon 5 (Fig. 1). Therefore, this study examined whether the white plumage phenotype in the black Yeonsan Ogye population is due to this change in the TYR gene.
MATERIALS AND METHODS
The study examined 19 of 625 Yeonsan Ogye chickens from the same generation: seven white, four brindle, and eight normal black chickens (Fig. 2). Genomic DNA (gDNA) was extracted from blood collected from the brachial vein. After mixing 20 μL of blood with 180 μL of phosphate-buffered saline, gDNA was extracted using the PrimePrep Genomic DNA Extraction Kit from Blood (GeNetBio, Daejeon, Korea), following the manufacturer’s instructions. The purity and concentration of the extracted DNA were confirmed using a NanoDrop 2000c spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The final gDNA was diluted to a concentration of 25 ng/μL and stored at -20°C before use.

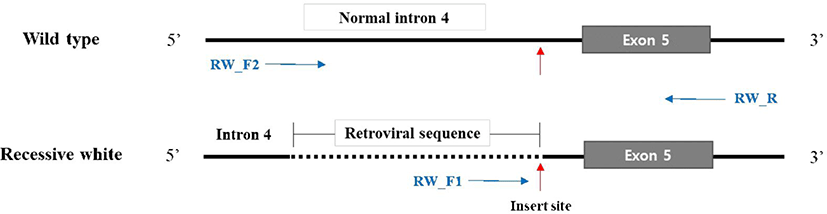
A primer set consisting of two forward primers and one reverse primer was prepared to identify the recessive white genotype based on published information (Fig. 3 and Table 1) [12]. Polymerase chain reaction (PCR) was performed in a total volume of 20 μL, which consisted of 2 μL of gDNA (25 ng/μL), 10 μL of HS Prime Taq Premix (2×) (GeNetBio), 0.5 μL each of the three primers (20 pmol/μL), and distilled water to make up the remaining volume. PCR consisted of an initial 3 min at 95°C, followed by 35 cycles of 95°C for 30 s, 58°C for 30 s, and 72°C for 1 min, and a final 5 min extension at 72°C. The product was examined via electrophoresis at 130V for 40 min using 2% agarose gels.
The PCR products were sequenced and compared with sequences of the TYR gene (NCBI accession: NC_006088.5) and intron 4 of the TYR gene with the ev-1 insertion (GenBank: DQ118 701.1) published in the NCBI database (https://www.ncbi.nlm.nih.gov). Another primer set with two forward and two reverse primers was produced from these sequences using Primer-BLAST on the NCBI website (Table 1). PCR amplification was performed at 95°C for 3 min, followed by 35 cycles of 95°C for 30 s, 61°C for 30 s, and 72°C for 1 min, and a final 5 min extension at 72°C. The amplified DNA fragments were purified with a PrimePrep PCR Purification Kit (GeNetBio) and sequenced using an automatic genetic analysis machine (ABI3730XL, Applied Biosystems, Foster City, CA, USA).
RESULTS AND DISCUSSION
Only a few chicken breeds are entirely black like the Yeonsan Ogye. These include Ayam Cemani from Indonesia, H’Mong from Vietnam, Black Silkie from China, and Svarthona from Sweden [13,14]. In Asia, the nutritional value of black-bone chickens has long been known. Many studies have reported on the antioxidant and anti-inflammatory effects of immuno-active substances in black-bone chickens, such as carnosine and cytokine [15–19]. The Yeonsan Ogye is functional medicinal food. In the classical medical books “Dongeuibogam” by Heo and “Bonchogangmok” by Lee, the Yeonsan Ogye was considered effective for treating stroke, arteriosclerosis, and heart disease [20,21].
To maintain the Yeonsan Ogye population, the Yeonsan Ogye Foundation is cooperating with various livestock research institutes and universities [5]. However, there is no established system for selecting which males and females to breed other than selecting completely black individuals. Consequently, individuals with exceptional phenotypes continue to appear in the population.
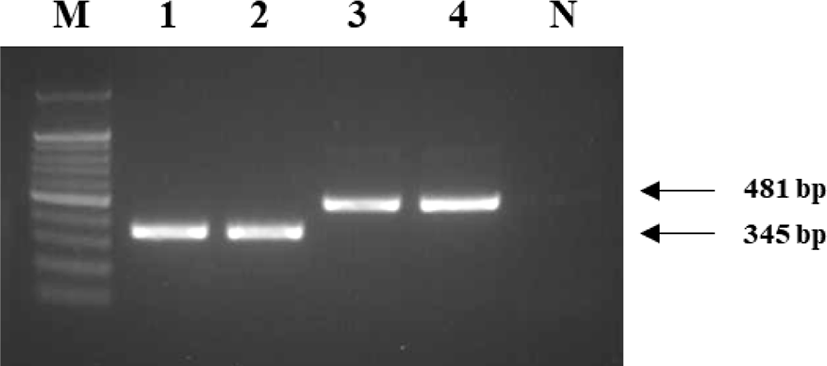
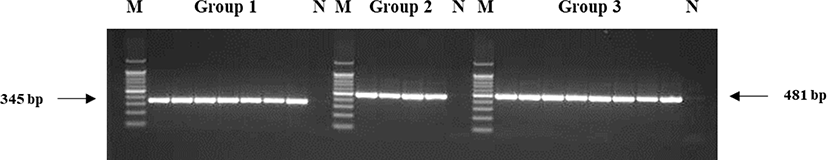
This study genotyped individuals with plumage that was not completely black, but either completely or partially white. PCR amplification products were confirmed using gel electrophoresis. Homozygous colored and recessive white individuals would produce a single 481 bp band on amplification with the primers RW_F2 and RW_R or a 345 bp band with the primers RW_F1 and RW_R, whereas heterozygous individuals would produce both bands (Fig. 4). The seven white chickens tested all produced one 345 bp band and were homozygous recessive white individuals. The four brindle and eight normal black chickens all produced one 481 bp band and were homozygous colored individuals. None of the individuals exhibited heterozygous patterns (Fig. 5).
The genotypes were confirmed by sequencing. On aligning the sequences with reference sequences (NCBI accession: NC_006088.5 and GenBank: DQ118701.1), the white chickens had the same sequence as the recessive white type (GenBank: DQ118701.1). Similarly, the sequences of the normal and brindle chickens matched that of the wild type (Fig. 6).
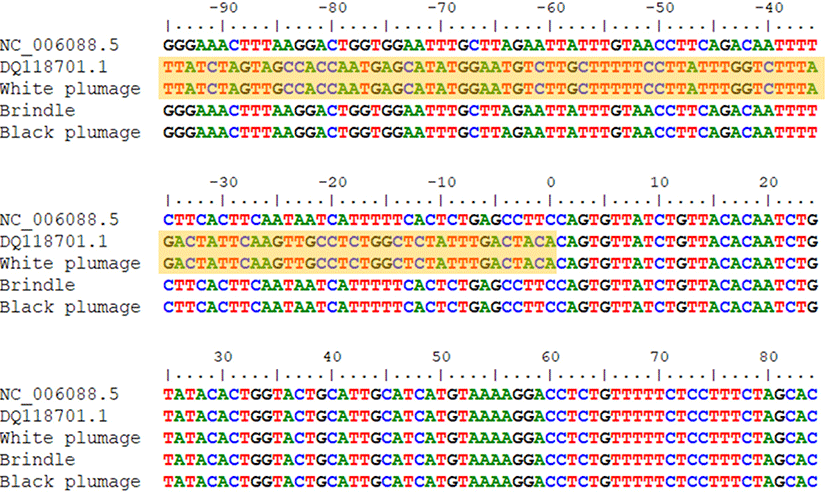
Comparing the white chicken sequences against the NCBI database using Nucleotide BLAST (BLASTN), the inserted retrovirus sequence exhibited 99% (278/279) identity (GenBank: DQ118701.1), as reported previously, and 98% (194/198) identity with the avian leukosis endogenous virus ev-1 sequence (GenBank: AY013303). The other sequences were the same as the TYR mRNA reference sequence (NCBI accession: NM_204160). Therefore, part of the retroviral sequence was inserted in the normal TYR intron 4. Retroviral insertion causes phenotypic changes by affecting the host genetic mechanism [22,23]. Specifically, this can be involved in pigmentation and control the expression of coat color [24,25]. The KIT gene is a proto-oncogene that causes cancer, but when a mutation occurs, it induces partial piebaldism [26,27]. The retroviral insertion identified here is located in TYR intron 4 and affects the expression of exon 5, a membrane-spanning domain that plays an important role in determining the location of melanosome enzymes [28]. Retroviral insertion interfered with the transcription of exon 5, resulting in impeded melanogenesis.
The exceptional phenotypes observed in the Yeonsan Ogye population include the recessive white mutation and a few individuals with a mixed plumage phenotype including yellow and gold coloration, similar to a brindle chicken. According to Yu et al. [29], four homeobox genes (HOXB9, HOXC8, HOXA9, and HOXC9), two glutathione metabolism genes (CHAC1 and GPX3), and the transforming growth factor beta signaling pathway are associated with plumage melanogenesis in black-bone chickens. Campo and Alvarez [30] found that the birchen allele (ER) at the E locus is affected by the melanotic gene (Ml) in the development of black plumage. Kerje et al. [31] reported that the extended black (E) locus controls plumage color in the same way as MC1R. Based on these results, an investigation of plumage color in Yeonsan Ogye can help with establishing a selection system to maintain its unique phenotype. It may also elucidate the reasons for the medicinal use of black Yeonsan Ogye.
As explained above, the Yeonsan Ogye has value as a genetic resource. But because only black individuals are selected to breed and there is no systematic breeding method, pedigree management has been poor. Additionally, because the individuals are selected phenotypically, it is impossible to confirm the presence or absence of mutant alleles. Therefore, to preserve the unique characteristics of the Yeonsan Ogye, genetic studies of specific phenotypes are required, and a new selection system for maintaining genetic diversity must be established.
CONCLUSION
This study genotyped Yeonsan Ogye chickens and confirmed that white chickens had the recessive white (c) TYR allele. Sequencing showed that an ev-1 retroviral sequence was inserted at the end of TYR intron 4 in this allele. Since the size of the sample used in this study was limited, further research through securing additional samples to perform validation studies is necessary. Therefore, after validation studies, a selection system for conserving the phenotypic characteristics and genetic diversity of the population could be established if additional studies to elucidate specific phenotype-related genes in Yeonsan Ogye are performed.