INTRODUCTION
Mesenchymal stem cells (MSCs) are multipotent stromal cells recognized in regenerative medicine and transplantation for their potential to differentiate into various cell types and modulate immune responses [1–3]. They have been widely studied in several species, particularly humans and they have shown therapeutic potential to treat various conditions such as cardiovascular diseases, neurodegenerative disorders, and immune-mediated diseases [4,5]. In addition to humans, studies have also been conducted on the characteristics of MSCs from various animal species, including dogs, goats, pigs, rabbits, and sheep, which generally exhibit positive CD44 expression and negative CD45 expression [6–9]. Furthermore, recent reports highlight the application of MSCs in treating conditions such as musculoskeletal diseases, skin disorders, ocular diseases, neuromuscular disorders, chronic gingivitis, inflammatory bowel disease, and asthma in companion animals [10–12]. However, the biological characteristics and potential applications of porcine MSCs remain unclear and require further investigation [5]. Pigs are known to share significant similarities with humans in physical, biochemical, anatomical, and gene expression patterns, making them valuable as preclinical trial animals [13–15]. Moreover, the high functional and anatomical similarity of the heart and kidney to those of humans has led to the recent use of pigs as a means for xenotransplantation [16–19]. Consequently, porcine MSCs are particularly valuable for preclinical research and therapeutic applications, including their role in xenotransplantation [20].
The application of porcine bone marrow (BM)-derived MSCs (BM-MSCs) is promising in veterinary medicine and as a model for studying disease mechanisms and developing therapeutic strategies in translational research [21]. Their immunomodulatory properties suggest that they may be crucial to reducing immune responses associated with graft rejection, making them a promising tool for improving the success of organ and tissue transplantation [22–25]. Notably, their ability to promote tissue repair and reduce inflammation has been reported in recent studies, reinforcing their potential use in regenerative medicine [26–29].
MSCs are typically identified by specific surface markers critical for their immunomodulatory functions, such as CD73, CD90, and CD105; however, they lack hematopoietic markers, such as CD45 [6,21,30]. These markers have been extensively used to characterize MSCs across different species, providing a basis for their identification and therapeutic applications [31,32]. Despite the recognized importance of these markers, limited data exist on the expression profiles and functional characteristics of porcine BM-MSCs [33]. A comprehensive understanding of these characteristics is crucial for developing effective MSC-based therapies and enhancing transplantation success [34,35].
In the present study, we aimed to profile the characteristics of porcine BM-MSCs by comparing their expression patterns with those of porcine peripheral blood mononuclear cells (PBMCs), BM-attached cells (BMACs), and the porcine kidney epithelial cell line (PK(15)). BMACs and PBMCs were chosen as comparators because they represent distinct populations within the bone marrow and peripheral blood compartments. BMACs, which include stromal cells, macrophages, and other bone marrow-derived cells, support stem cell function [36,37], while PBMCs are peripheral immune cells used to understand the immunomodulatory properties of BM-MSCs [38,39]. This comparison highlights the regulatory mechanisms of BM-MSCs and provides insights into their therapeutic potential in regenerative medicine and transplantation. Using quantitative real-time polymerase chain reaction (qRT-PCR), flow cytometry, immunocytochemistry, and RNA sequencing, we aimed to elucidate the molecular and phenotypic features that distinguish BM-MSCs from other cell types. This study focused on the immunomodulatory functions and potential applications of BM-MSCs in mitigating graft rejection and promoting tissue regeneration.
MATERIALS AND METHODS
Porcine BM-MSCs (Cell Biologics), purchased from Cell Biologics and isolated from porcine tibias and femurs, were cultured in mesenchymal cell medium (Cell Biologics) containing 10% heat-inactivated fetal bovine serum (FBS, Gibco) and 1% penicillin/streptomycin (P/S, Cell Biologics) at 37°C in an incubator with a 5% carbon dioxide (CO2) atmosphere. Porcine BM cells were isolated from the humerus, tibia, and femurs of stillborn piglets. After a 10-d culture period in a culture dish, non-adherent cells were removed by discarding the supernatant. The remaining adherent cells were cultured and indicated as BMACs. BMACs and PK(15) (American Type Culture Collection) cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco) containing 10% FBS, 1% minimum essential medium non-essential amino acid solution, and 1% P/S at 37°C in an incubator with a 5% CO2 atmosphere.
THP-1 cell line (Korean Cell Line Bank) was cultured in Roswell Park Memorial Institute 1640 medium (RPMI 1650, Gibco) containing 10% FBS and 1% P/S (Gibco) at 37°C in an incubator with a 5% CO2 atmosphere. To obtain phorbol-12-myristate-13-acetate (PMA)-differentiated THP-1 cells, THP-1 cells were differentiated using 10 ng/mL PMA (Sigma-Aldrich), and the PMA-free medium was changed the next day for 24 h. No contamination was detected in any cell cultures.
Total RNA was isolated using Trizol (Life Technologies). Total cellular RNA was used to synthesize complementary DNA (cDNA) using a QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer’s instructions.
Quantitative PCR (qPCR, Power SYBRTM Green PCR Master Mix, 4368702, Applied Biosystems) was performed using porcine primers for glyceraldehyde-3-phosphate dehydrogenase (GAPDH), CD73, CD90, and CD105 and human primers for GAPDH, tumor necrosis factor-alpha (TNFα), interleukin (IL)-6, IL-10, C-C chemokine receptor type 7 (CCR7), and CD163. All qPCR primers were designed using Primer 3V0.4.0 (Table 1). qPCR was performed as follows: 95°C for 10 min, 40 cycles at 95°C for 15 s, and 60°C for 1 min on a PCR machine (A28134, Applied Biosystems). Messenger RNA (mRNA) levels were determined using GAPDH (△Ct = Ct gene of interest –Ct GAPDH) and reported as relative mRNA expression (△△Ct = 2△Ct sample−△Ct control) or the fold change.
Cells in each group were collected in fluorescence-activated cell sorting tubes (BD Biosciences) and washed twice with ice-cold phosphate-buffered saline (PBS). BM-MSCs and PBMCs were stained with allophycocyanin (APC)-conjugated CD44 (Abcam), CD45 (Bio-Rad), CD73 (Invitrogen), CD90 (Abcam), and CD105 (Invitrogen) for 1 h at room temperature. Alexa Fluor 488-conjugated goat anti-mouse immunoglobulin (Ig) G (Invitrogen), 488-conjugated donkey anti-sheep IgG (Invitrogen), and 568-conjugated goat anti-mouse IgG (Invitrogen) were used for cell labeling. Stained cells were analyzed using flow cytometry (Beckman Coulter) and CytExpert software (Beckman Coulter). For each sample, a cell count of 5,000 cells was obtained. The region of each sample was selected for the forward and side scatters, and a histogram was used to measure the mean fluorescence intensity of fluorescein isothiocyanate, phycoerythrin, or APC.
After fixation in 4% paraformaldehyde in Dulbecco’s PBS, the cells were stained with CD45 (Bio-Rad, MCA1222GA), CD73 (Invitrogen), and CD105 (Invitrogen). Alexa Fluor 647-conjugated goat anti-mouse IgG (Invitrogen) and 488-conjugated donkey anti-sheep IgG (Invitrogen) were used for cell labeling. Nuclei were stained with a mounting medium containing 4’,6-diamidino-2-phenylindole (Abcam). A confocal microscope (ZEISS) was used to obtain images.
Notably, 1 μg RNA was isolated from 3 × 106 cells using the phenol/chloroform extraction method. RNA integrity was assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies). Each cDNA library was prepared using a QuantSeq 3mRNA-seq Library Prep Kit (Lexogen). The entire process, including sequencing, mapping, and normalization, was performed according to the manufacturer’s instructions. Differentially expressed genes (DEGs) were determined from the genes with expression levels changed as |Log2 (fold change)| ≥ 2 (Supplementary file 1). Excel-based DEG Analysis (ExDEGA; E-biogen) was used to visualize the hierarchical heatmap and create a Venn diagram of DEGs.
To compare functional annotations among BM-MSCs, BMACs, and PBMCs, Kyoto Encyclopedia of Genes and Genomes pathway analysis was performed using the Database for Annotation, Visualization, and Integrated Discovery Bioinformatics Resources 6.8 (Supplementary file 2) [40, 41]. Furthermore, upstream regulators, such as the main biological network, canonical pathway, and upstream regulator identification, were analyzed using IPA (Qiagen).
PMA-differentiated THP-1 cells were treated with 1 µg/mL lipopolysaccharide (LPS; Sigma-Aldrich, LPS25) for 24 h and seeded in a 12-well plate (Greiner Bio-One, 665180) at a density of 1×106 cells/well. The same number of BM-MSCs was seeded in Transwell inserts (Greiner Bio-One, 665640). After 24 h, PMA-differentiated THP-1 cells in the bottom plate were evaluated for IL-1β, IL-6, TNF𝛼, IL-10, CCR7, and CD163 mRNA expression using qRT-PCR.
All data are presented as the mean ± SD. All experiments were performed at least thrice. Statistical significance was determined using Student’s t-test (two-tailed) or analysis of variance using GraphPad Prism 8 software (GraphPad). The p-value and Z-score were calculated using the computational algorithms of Student’s t-test and Fisher’s exact test to confirm statistical significance.
RESULTS
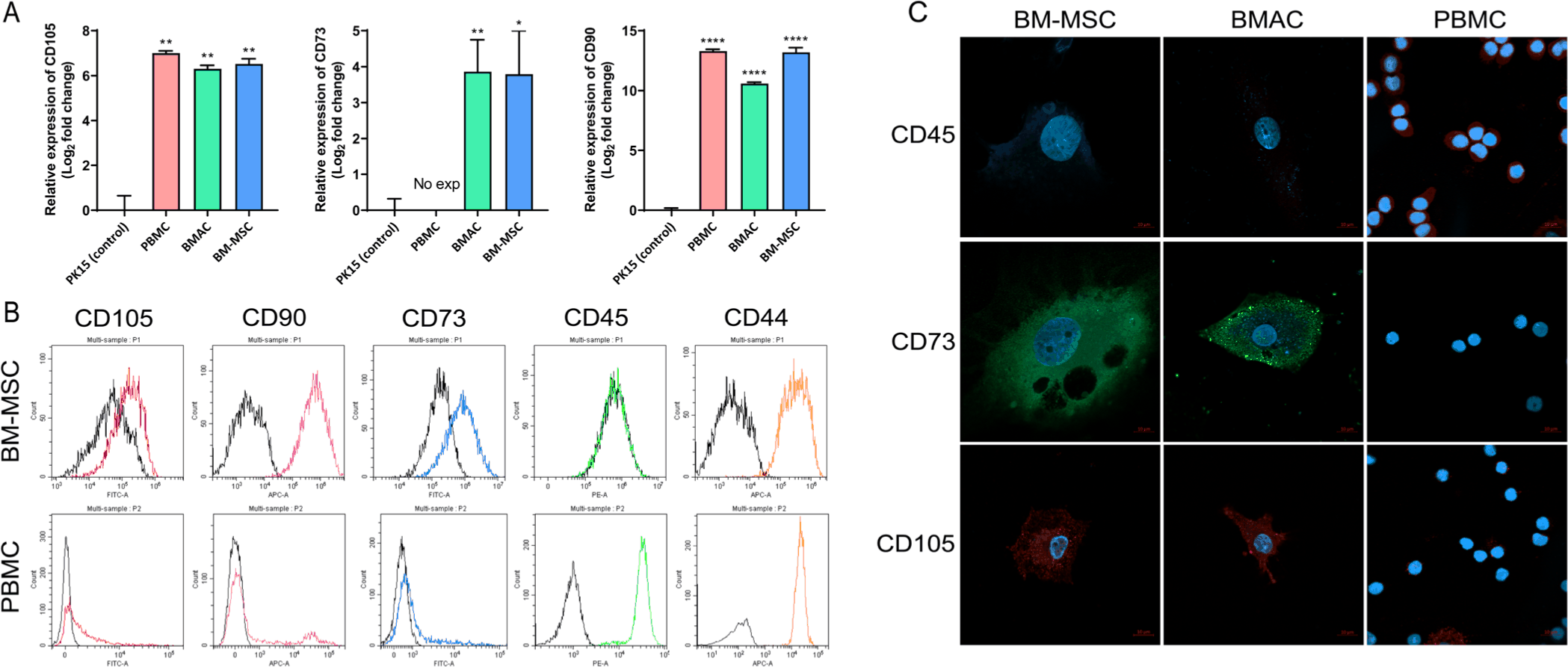
To characterize porcine BM-MSCs, BM-MSCs obtained from Cell Biologics were compared with porcine PBMCs, BMACs, and PK(15) cells to analyze the expression patterns of BM-MSC markers. Previous studies have reported that MSCs can be identified and characterized based on the expression of specific surface markers. First, we analyzed the mRNA expression patterns of CD105, CD73, and CD90, which are human MSC markers, in BM-MSCs, PBMCs, and BMACs and compared them to those in PK(15) cells using qRT-PCR. The mRNA expression levels of CD73, CD90, and CD105 were confirmed in BM-MSCs and BMACs and exhibited an expression pattern consistent with that observed in human MSCs (Fig. 1A). Using flow cytometry, BM-MSCs demonstrated strong positive expression for CD44, CD73, CD90, and CD105, while showing negative expression of CD45, confirming their mesenchymal identity (Fig. 1B). In contrast, PBMCs, composed of a heterogeneous cell type, generally express all markers, with CD44 and CD45 being universally expressed across all cells (Fig. 1B). Immunocytochemistry further revealed that BM-MSCs were negative for CD45 and positive for CD73 and CD105, which is consistent with the results observed for BMACs, whereas PBMCs showed all-positive expression for CD45, while the expression of CD73 and CD105 was barely detected (Fig. 1C). These findings indicate that porcine BM-MSCs maintain a distinct MSC marker expression, which clearly differentiates them from PBMCs.
To analyze the differential gene expression patterns among porcine BMACs, BM-MSCs, and PBMCs, we performed a comprehensive gene expression analysis using mRNA-Seq data. The DEGs between BMACs and PBMCs and between BM-MSCs and PBMCs were compared (Fig. 2A). As shown on the Venn diagram, 1,297 upregulated and 1,399 downregulated genes were observed in the comparison between BMACs and PBMCs, whereas 1,873 upregulated and 2,062 downregulated genes were observed in the comparison between BM-MSCs and PBMCs. The overlap included 4,467 upregulated and 4,798 downregulated genes, with 365 contra-regulated genes shared between comparisons.
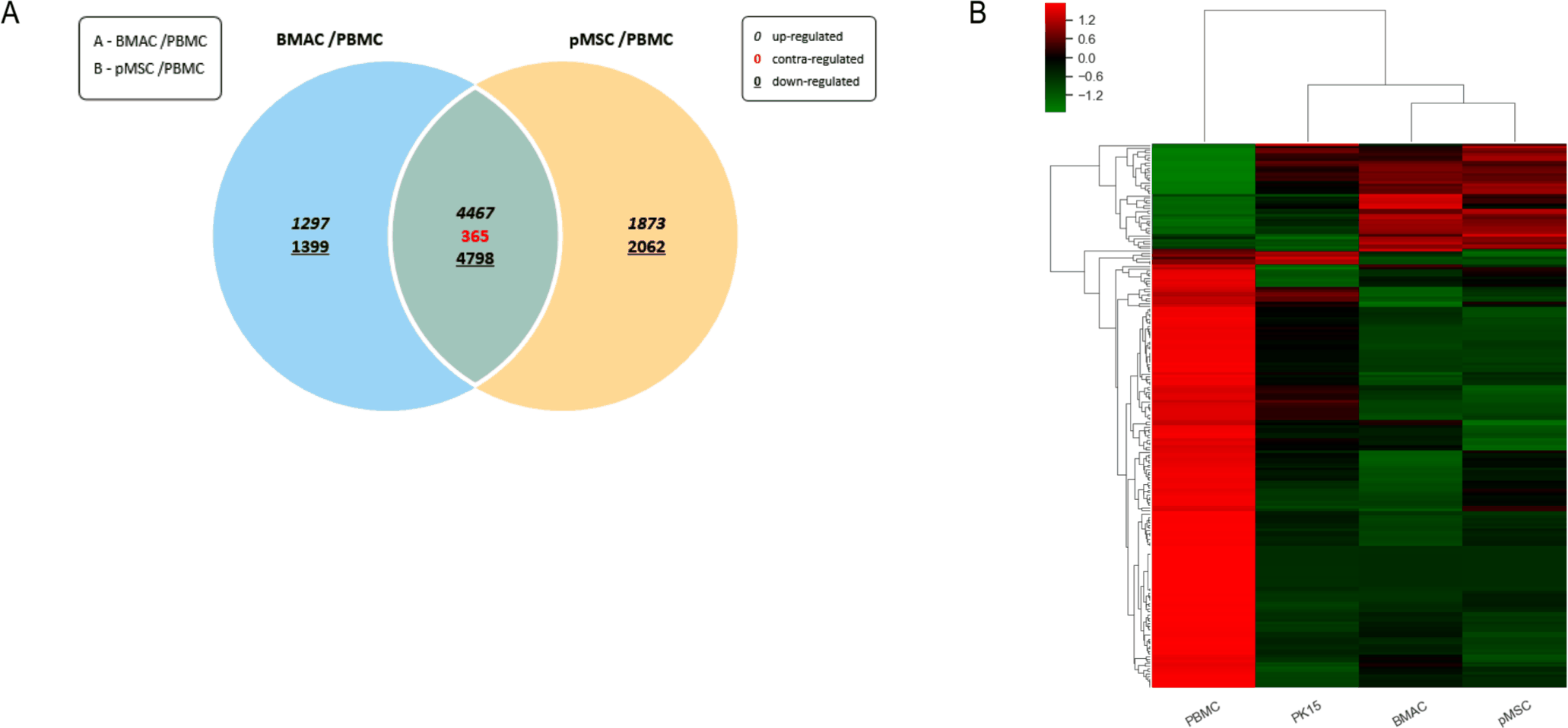
Gene expression profiles were visualized using a clustering heatmap (Fig. 2B), which showed the hierarchical clustering of gene expression profiles across PBMCs, PK(15) cells, BMACs, and BM-MSCs. The clustering revealed distinct gene expression profiles, highlighting the unique regulatory mechanisms of each cell type. These results indicate that BM-MSCs exhibit a different expression pattern from that of PBMCs but a significantly similar expression pattern to that of BMACs.
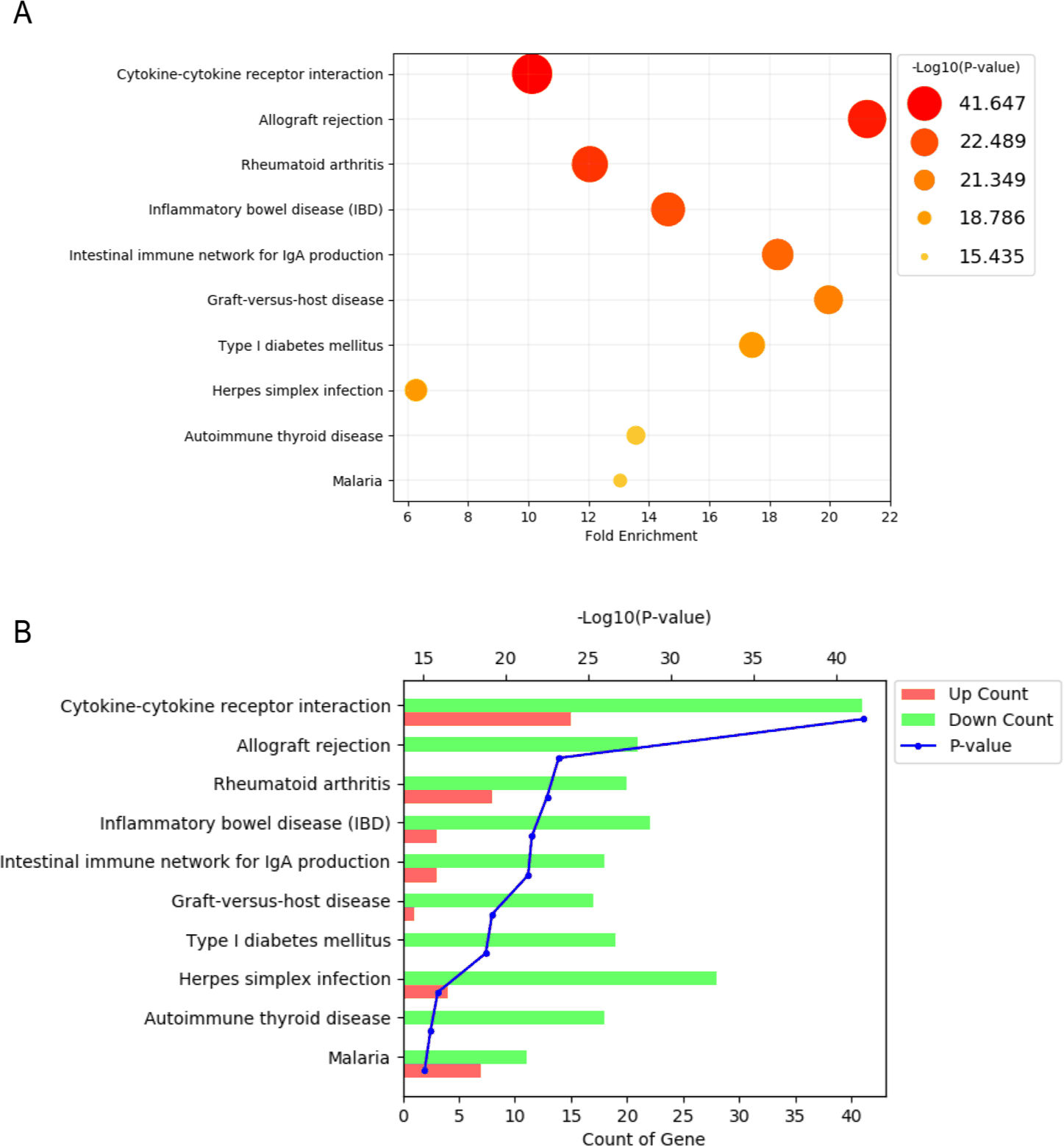
To identify significant biological pathways associated with BM-MSCs, gene ontology (GO) enrichment analysis of DEGs was performed using the ingenuity pathway analysis (IPA) software. In this analysis, DEGs were subjected to pathway enrichment analysis to identify significant changes in BM-MSCs compared with those in PBMCs and BMACs. The statistical significance (p-value) of each pathway was determined, and pathways with a p-value of ≤ 0.05 were considered significant.
The significant pathways identified by GO analysis are shown in Fig. 3. The gene enrichment assay revealed the most enriched pathways, with each bubble representing one pathway (Fig. 3A). The size and color of the bubble indicate the fold enrichment and significance level, respectively. The key pathways identified were cytokine-cytokine receptor interaction, allograft rejection, rheumatoid arthritis, inflammatory bowel disease, and the intestinal immune network for IgA production. Furthermore, as shown in Fig. 3B, the pathway enrichment bar plot shows the number of upregulated (red) and downregulated (green) genes for each significantly enriched pathway, with the blue line indicating the p-value. This plot further shows the significant pathways identified in the pathway enrichment bubble plot.
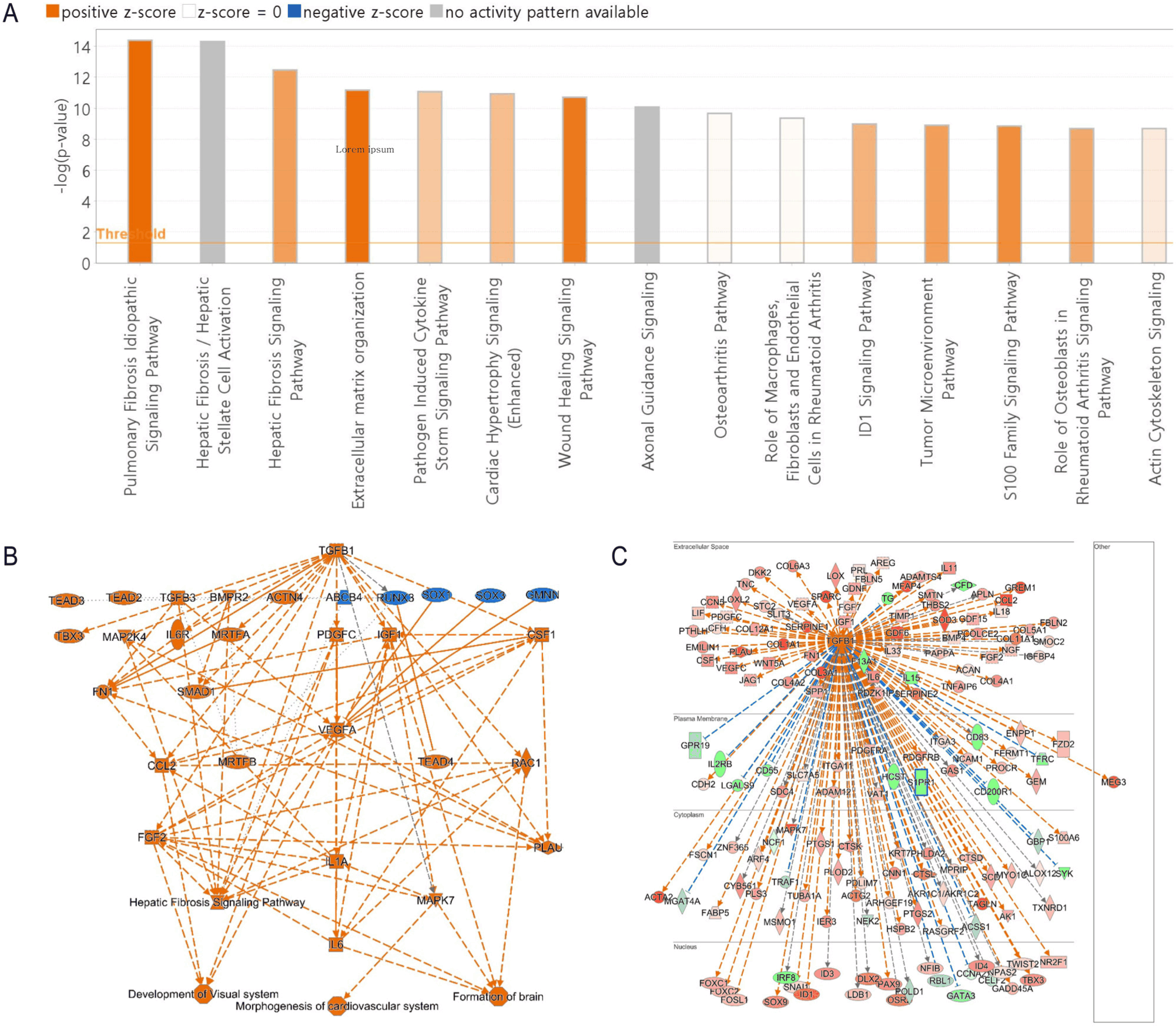
Using IPA, we performed a pathway enrichment analysis of DEGs identified in porcine BM-MSCs compared with those in PBMCs. This analysis revealed several key canonical pathways, with significant z-scores indicating either activation or inhibition. The top biological functions were the pulmonary fibrosis idiopathic signaling pathway, hepatic fibrosis/hepatic stellate cell activation, hepatic fibrosis signaling pathway, extracellular matrix organization, and the pathogen-induced cytokine storm signaling pathway (Fig. 4A). The network analysis of DEGs in porcine BM-MSCs revealed the central role of the transforming growth factor-beta (TGF-β) signaling pathway, linking key downstream pathways involved in cellular differentiation, fibrosis, and immune response modulation (Fig. 4B). Furthermore, the biological network of TGF-β as an upstream regulator in the subcellular environment indicates the extensive regulatory influence of TGF-β on a wide array of genes associated with tissue repair, immune modulation, and cellular homeostasis (Fig. 4C). These findings collectively emphasize the intricate signaling networks active in BM-MSCs, highlighting the significant role of TGF-β in immune regulation.
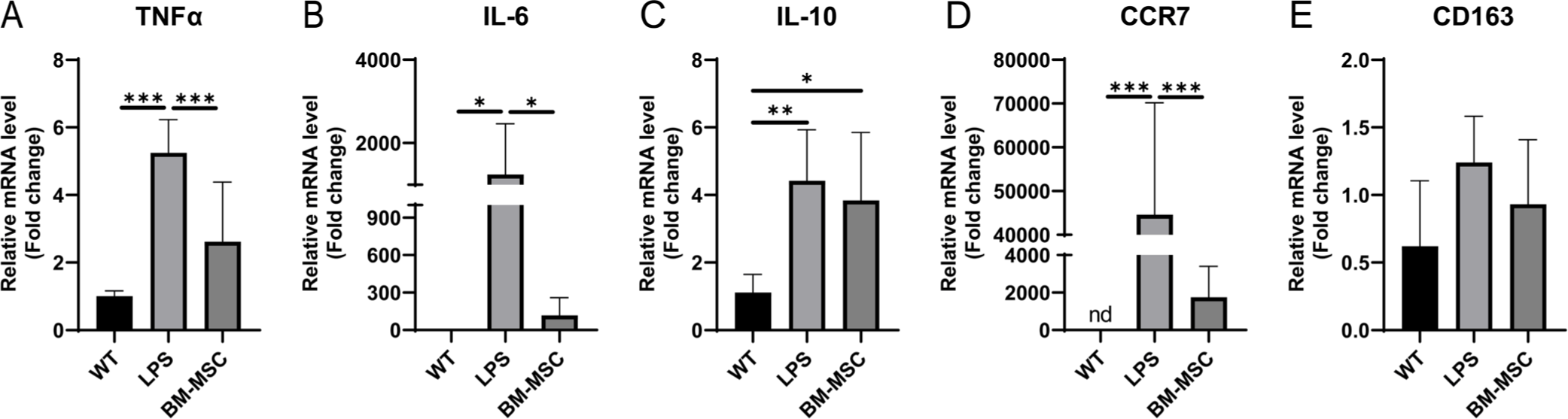
Our data revealed that BM-MSCs exhibited higher TGF-β expression levels than PBMCs. To evaluate the immunomodulatory effects of BM-MSCs under xenogeneic conditions, PMA-differentiated THP-1 cells, treated with 1 µg/mL LPS for 24 h, were indirectly co-cultured with BM-MSCs using a Transwell system. The expression levels of key cytokines and markers associated with inflammation were also assessed. The results revealed a significant decrease in the expression of pro-inflammatory cytokines, IL-6 and TNFα, in PMA-differentiated THP-1 cells treated with LPS and co-cultured with BM-MSCs (BM-MSC group) compared with the LPS group (Figs. 5A and 5B). In contrast, the anti-inflammatory cytokine IL-10 was significantly upregulated in the BM-MSC group, and its mRNA levels were maintained (Fig. 5C). In addition, significant downregulation of the expression of CCR7, a marker associated with the M1 macrophage phenotype, and slight upregulation of the expression of CD163, a marker for the M2 macrophage phenotype, were observed in the BM-MSC group compared with the WT group (Figs. 5D and 5E). These findings suggest that BM-MSCs exert a potent immunomodulatory effect by suppressing pro-inflammatory responses and promoting an anti-inflammatory M2-like macrophage phenotype under xenogeneic conditions.
DISCUSSION
In the present study, we provided a detailed characterization of porcine BM-MSCs and compared their gene expression profiles and immunomodulatory properties with those of PBMCs and BMACs. Our findings offer significant insights into the molecular and phenotypic distinctiveness of BM-MSCs, emphasizing their potential for therapeutic applications in transplantation and regenerative medicine.
A key aspect of the present study was the use of complementary techniques, including qRT-PCR, flow cytometry, immunocytochemistry, and RNA sequencing. These comprehensive techniques enabled us to confirm that classical MSC markers, including CD44, CD73, CD90, and CD105, were expressed in BM-MSCs, whereas the hematopoietic marker, CD45, was not observed. This expression profile was consistent with the established criteria for MSC identification across different species, indicating the conserved nature of these markers [6,42–44]. It has been reported in some studies that CD73 and CD105 are not expressed in porcine BM-MSC, unlike their human counterparts [45,46]. However, our data confirmed the RNA and protein expression of these markers in porcine BM-MSCs, aligning them more closely with the characteristics of human BM-MSCs [47,48]. BMACs are a heterogeneous population of cells, including macrophages, stromal cells, and other bone marrow-derived cells, that provide a supportive environment for stem cell function [49,50]. The similarity in gene expression patterns between BM-MSCs and BMACs suggests that BM-MSCs retain their stem cell characteristics. Furthermore, the distinct expression patterns in BM-MSCs compared with those in PBMCs may enhance their therapeutic potential, particularly in tissue regeneration and immune modulation [51].
Transcriptome profiling revealed significant differences between BM-MSCs and PBMCs, with a significant number of DEGs observed (Fig. 2). This differential expression underscores the unique regulatory mechanisms inherent in BM-MSCs, which are potentially advantageous for regulating immune responses [52,53]. Notably, all of the upregulated genes in the top 10 DEGs are located downstream of the TGF-β signaling pathway, a finding further corroborated by the IPA analysis (Supplementary file 3, Figs. 4B and 4C). These results suggest that the differences in unique regulatory mechanisms between PBMCs and MSCs are primarily driven by the TGF-β pathway. Additionally, the overlap of upregulated and downregulated genes between BM-MSCs and BMACs suggests that both cell types share common regulatory pathways. GO enrichment analysis revealed key pathways significantly associated with BM-MSCs, such as cytokine-cytokine receptor interaction and allograft rejection. These pathways are crucial for modulating immune responses and promoting tissue repair, thereby highlighting the therapeutic potential of BM-MSCs for transplantation [54–56].
BM-MSCs exhibited higher TGF-β levels than PBMCs, indicating their central role in immune regulation and immunomodulatory functions [4,57,58]. Our pathway enrichment and network analyses revealed TGF-β signaling as a pivotal node that connects various downstream pathways involved in fibrosis, cellular differentiation, and immune regulation (Fig. 3). This finding is consistent with those of recent studies, emphasizing the importance of TGF-β in maintaining immune homeostasis and facilitating tissue repair [59,60]. Recent studies also indicate that TGF-β, produced by BM-MSCs, plays a role in influencing the proliferation of CD34+ cells and regulating hematopoiesis [61]. Furthermore, we observed a reduction in pro-inflammatory cytokines (IL-6 and TNFα) and an upregulation of the anti-inflammatory cytokine, IL-10, in the BM-MSC group under xenogeneic conditions [62]. Additionally, the expression of CCR7, an M1 macrophage marker, was significantly decreased, while CD163, an M2 macrophage marker, was increased in the BM-MSC group [63]. These results suggest that BM-MSCs regulate immune responses through downstream signals mediated by TGF-β, leading to the polarization of pro-inflammatory M1 macrophages into anti-inflammatory M2 macrophages under both allo-reactive and xenogeneic conditions.
These results are promising; however, certain challenges must be addressed before BM-MSCs can be widely applied in clinical settings. One significant issue is the long-term safety and efficacy of BM-MSC-based therapies, particularly in xenogeneic contexts where immune rejection remains a major concern [20]. TGF-β is an immunoregulatory cytokine that plays a crucial role in the differentiation of Th9, Th17, and regulatory T cells, and its influence has been extensively studied in both acute and chronic responses in allogeneic transplantation [64]. Also, TGF-β acts on macrophages to induce an anti-inflammatory response via the Smad2/3 pathway and promotes M2-like macrophage polarization [65,66]. A previous study has shown that BM-MSCs secreting TGF-β, when administered to septic mice, significantly reduced inflammatory macrophages, suggesting that TGF-β can regulate immune responses, at least during the acute phase [67]. Although our findings were obtained under xenogeneic conditions and in vitro, they exhibit a similar pattern (Fig. 5). Furthermore, the higher levels of TGF-β expression in BM-MSCs and their capacity to induce an anti-inflammatory macrophage response indicate their potential to reduce graft rejection and improve transplant outcomes [68,69]. However, further research is necessary to fully elucidate the mechanisms through which BM-MSCs exert these effects, particularly in long-term studies, and to assess the efficacy and safety of BM-MSC-based therapies in clinical settings.
In conclusion, our study provides a comprehensive profile of porcine BM-MSCs and describes their distinct molecular characteristics and immunomodulatory potential. Our findings support the ongoing investigation of BM-MSCs in the context of xenotransplantation and regenerative medicine with the aim of developing novel therapies that can effectively manage immune responses and enhance tissue regeneration.