INTRODUCTION
Oxidative stress is defined as a state of reactive oxygen species (ROS) imbalance, which attenuates the body’s antioxidant capacity [1]. In poultry, oxidative stress results from environmental stress factors, such as temperature, stocking density, and lighting [2]. Such stress factors can disrupt the antioxidant system balance in intestinal epithelial cells, causing lipid peroxidation, protein nitration, DNA damage, and apoptosis [3]. A damaged intestinal mucosa negatively impacts nutrient digestion and absorption in chickens [4]. Therefore, functional nutrients may mitigate oxidative stress in chicken intestinal epithelial cells (cIECs) need to be identified. Amino acids, vitamins, and minerals play important roles (e.g., metabolism, physiology, and immunity) in the intestinal mucosa [5,6].
Threonine (Thr) serves a vital role in protein synthesis and maintenance by synthesizing intestinal mucin [7,8]. After being absorbed in the intestines, it protects the intestines against pathogens and anti-nutritional factors and helps them function properly via oxidized mucosal cells [9,10]. In birds, arginine (Arg) lacks the enzymes involved in its synthesis, necessitating dietary supplementation, unlike that in mammals [11]. It is an essential nutrient for tissue healing and a critical component of immune regulation. Additionally, it is a key amino acid that promotes growth hormone secretion and serves as a precursor for polyamine synthesis, which is requisite to intestinal healing [12–14]. Vitamin C acts as an antioxidant by donating electrons to free radicals and ROS and inhibiting lipid peroxidation. In addition, it maintains intestinal absorption function and alleviates the oxidative stress-induced decrease in nutrient absorption [15]. Vitamin E is a fat-soluble antioxidant that protects lipoproteins and cell membranes. It improves total antioxidant capacity and decreases oxidative stress and immune indicators (e.g., interleukin-6 and tumor necrosis factor-alpha) [16]. Chromium (Cr) plays a crucial role in enhancing glucose uptake within cells by augmenting insulin receptor beta-kinase activity [17]. This mechanism promotes efficient glucose transportation and allays oxidative stress [18]. Furthermore, Cr stimulates antioxidant enzyme synthesis, combats lipid peroxidation, modulates nuclear factor erythroid 2-related factor 2 expression, activates the AMP-activated protein kinase pathway, and inhibits the mitogen-activated protein kinase pathway [19]. Zinc (Zn) induces metallothionein, which synthesizes copper–zinc superoxide dismutase (Cu–Zn SOD), an antioxidant enzyme. It also exhibits excellent binding ability to metal ions. Moreover, Zn helps increase glutathione (GSH) levels and reduces iron-induced oxidative stress in the intestines. In addition, it maintains the stability of the sulfhydryl group, which has strong reducing power [20]. Qiu et al. [21] and Wang et al. [22] found Thr and Arg supplementation to induce significantly greater cell viability in porcine intestinal epithelial cells (IPEC-J2). Miller et al. [23] reported that vitamin C can reduce apoptosis and necrosis in human gastric epithelial cells. Vitamin E supplementation reportedly elicits significantly greater GSH and less thiobarbituric acid-reactive substance levels in rat intestinal epithelial cells [24]. Kim et al. [25] revealed that Cr significantly increases cell viability in oxidative stress-induced cIECs. Kilari et al. [26] demonstrated that Zn can significantly increase metallothionein levels and prevent intracellular GSH depletion in human intestinal epithelial cells (Caco-2). Therefore, six functional nutrients may alleviate oxidative stress in cIECs by improving cell viability and antioxidant enzyme production.
In vitro experiments serve an indispensable role in resolving ethical issues before performing in vivo experiments. However, most in vitro studies focus on human and rat intestinal epithelial cells. Studies investigating the effects of functional substances on cIECs are lacking. Discovering substances may mitigate oxidative stress in cIECs is imperative.
Therefore, the current experiment endeavored to investigate the protective effect of functional nutrients on oxidative stress-induced cIECs.
MATERIALS AND METHODS
The cIECs were isolated and cultivated according to the method described by Kim et al. [25], with slight modifications. Eggs were purchased from specific pathogen-free-birds (VALO BioMedia GmbH) and incubated using an egg-hatching incubator. Primary cells were isolated from chick embryos at 16 days of embryogenesis. The intestine was washed with phosphate-buffered saline to remove blood and impurities and cut into small fragments (0.5–1 cm) using a sterile scalpel blade. Thereafter, the fragments were weighed and placed in a 50-mL tube along with digestive enzymes. The sample-to-digestive enzyme ratio was as follows: 1 g of tissue to 1 mL of 2 U collagenase D, 1 mL of 4 U Dispase® II, and 50 μL of 100 mM CaCl2 (final concentrations: 1 U/mL collagenase D, 2 U/mL Dispase® II, and 2.5 mM CaCl2, respectively). The intestinal fragments were digested at 37°C for 2 hours and subsequently filtered through cell strainers (40, 70, and 100 μm) to isolate crypts from larger intestinal cells. The isolated crypts were centrifuged at 1,561×g for 3 minutes. Thereafter, the supernatant was discarded, and the cell pellet was washed with washing medium (Dulbecco’s modified Eagle medium [DMEM], 2% D-sorbitol, 2.5% fetal bovine serum [FBS], 100 μg/mL penicillin/streptomycin, and 2,500 μg/mL gentamicin) and subsequently centrifuged at 1,561×g for 10 minutes to isolate cIECs. The cIECs were treated with 10 mL of growth medium (DMEM, 2.5% FBS, 10 μg/mL insulin, 100 μg/mL penicillin/streptomycin, 2,500 μg/mL gentamicin, and 1,400 μg/mL hydrocortisone). Afterward, the cIECs were cultured in a cell dish coated with fibronectin and incubated at 37°C for 24 hours in 5% CO2. The cells were cultured in culture medium (DMEM, 2.5% FBS, 10 μg/mL insulin, and 100 μg/mL penicillin/streptomycin) every 2–3 days at 37°C for 24 hours in 5% CO2.
Cells were seeded into 96-well plates at a density of 1 × 104 cells/well and incubated at 37°C for 24 hours in 5% CO2. The experimental procedure followed the method of Kim et al. [25]. Briefly, each treatment group was treated with 2, 20, and 200 μg/mL of Thr, Arg, vitamin C, vitamin E, Cr, or Zn. After 24 hours, all wells, except the positive control (PC), were treated with 20 μM lipopolysaccharide (LPS) for 4 hours. Thereafter, cell viability was measured using the EZ-Cytox assay (DoGenBio), following the manufacturer’s protocol. After incubation, 20 μL of EZ-Cytox assay solution was added to each well, and the cells were subsequently incubated for 3 hours. Thereafter, the absorbance of the cells was determined at 450 nm using a microplate reader (INNO-S, Bio Mart).
Cells were seeded into 96-well plates at a density of 5 × 104 cells/well and incubated at 37°C for 24 hours in 5% CO2. Briefly, the treatment groups were treated with 2, 20, and 200 μg/mL. After 24 hours, all wells, except those of the PC, were treated with 20 μM LPS for 4 hours. Afterward, 50 μL of supernatant was transferred to a new 96-well plate. Finally, an equal volume of Griess reagent (1% sulfanilamide and 0.1% N-(1-Naphthyl)ethylenediamine in 5% HPO3) was immediately added to each well to measure the nitrite content at 540 nm.
The HSP70 was determined using a Chicken HSP70 ELISA Kit (Assay Genie), following the manufacturer’s protocol. Cells were seeded into 96-well plates at 1 × 104 cells/well density and incubated at 37°C for 24 hours in 5% CO2. Briefly, the treatment groups were treated with 2, 20, and 200 μg/mL of Thr, Arg, vitamin C, vitamin E, Cr, or Zn. After 24 hours, all wells, except the PC, were treated with 20 μM of LPS for 4 hours. Thereafter, the cell supernatant was transferred to a 96-well plate using a pipette and subsequently centrifuged at 4°C for 20 minutes at 1,561×g. The coated plate was washed twice with wash buffer, and 100 μL of each diluted and prepared standard, sample, and control (zero well) was placed in the designated well and incubated at 37°C for 90 minutes. The samples were discarded and washed twice. Thereafter, 100 μL of biotin-detection antibody working solution was added to the above wells and incubated at 37°C for 60 minutes. Afterward, the resulting mixture was washed thrice with wash buffer, and 0.1 mL of streptavidin–biotin complex working solution was added to each well. Subsequently, the plate was incubated at 37°C for 30 minutes under dark conditions. The incubated plate was washed 5 times, and 90 µL of 3,3’,5,5’-tetramethylbenzidine substrate was added to each well, followed by incubation at 37°C in the dark for 10–20 minutes. Thereafter, 50 µL of a stop solution was added to each well, and absorbance was measured at 450 nm using a microplate reader (INNO-S, Bio Mart) immediately after adding the stop solution.
The ROS level of LPS-induced cIECs was measured using 2’,7’-dichlorodihydrofluorescein diacetate (DCF-DA) dye [27]. cIECs were seeded into black 96-well plate at 1.0 × 104 cells/mL LPS (2 μg/mL) was added to the wells, and the plates were incubated for 4 hours at 37°C. Thereafter, DCF-DA solution (50 μM) was added to each well and incubated for 1 hour. The fluorescence of the 2’,7’-dichlorofluorescein product was measured at excitation and emission wavelengths of 480 and 530 nm using a microplate reader (INNO-S, Bio Mart).
The transepithelial electrical resistance (TEER) was measured using the method described by Chen et al. [28]. Briefly, differentiated cIECs in 0.4-μm pore apical chambers were treated with functional nutrients and LPS, as described above. Subsequently, 24-well plates were placed on a hot plate (Daihan Labtech) at 37°C. The TEER was measured at 0, 24, and 48 hours using an EVOM3 Epithelial Volt/Ohm Meter (World Precision Instruments), according to the manufacturer’s protocol.
Tight junction permeability in cIECs was measured using fluorescein isothiocyanate (FITC)–dextran, as demonstrated by Liu et al. [29]. Briefly, cells were seeded in the apical chamber of a 24-well Transwell® filter with 0.4-μm pores (SPL Life Sciences) at 1 × 104/well and incubated at 37°C for 24 hours in a humidified atmosphere containing 5% CO2. For cell differentiation, the culture medium was replaced with a differentiation medium containing 50 μg/mL dexamethasone (Sigma-Aldrich) every 2 days. On day 9 of the seeding process, the cells were incubated with the respective treatments for 24 hours and cotreated with or without 2 μg/mL LPS for 4 hours. After cell treatment, FITC–dextran (Sigma-Aldrich) dissolved in cell differentiation medium was added to the apical chamber at a final concentration of 2.2 mg/mL, and the well plates were incubated for 1 hour. Three 200-μL aliquots were taken from each well of a 24-well plate and added to a black 96-well plate (SPL Life Sciences). The amount of fluorescence in the black 96-well plate was measured using a microplate reader (INNO-S, Bio Mart) at excitation and emission wavelengths of 490 and 535 nm.
Statistical analyses were performed via analysis of variance using SPSS (SPSS, IBM), and differences among mean values were evaluated using Student–Newman–Keuls test. Each replicate was considered an experimental unit. The data were expressed as the mean ± standard deviation. Statistical significance was set at p < 0.05.
RESULTS
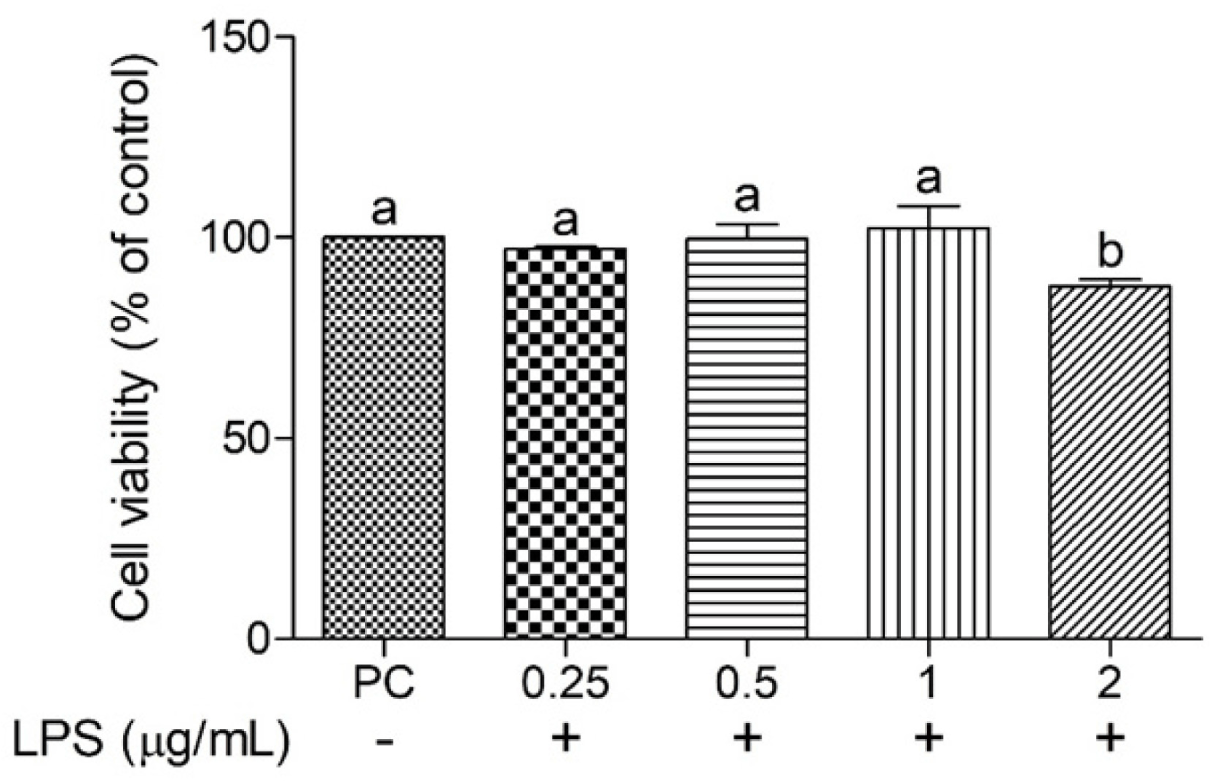
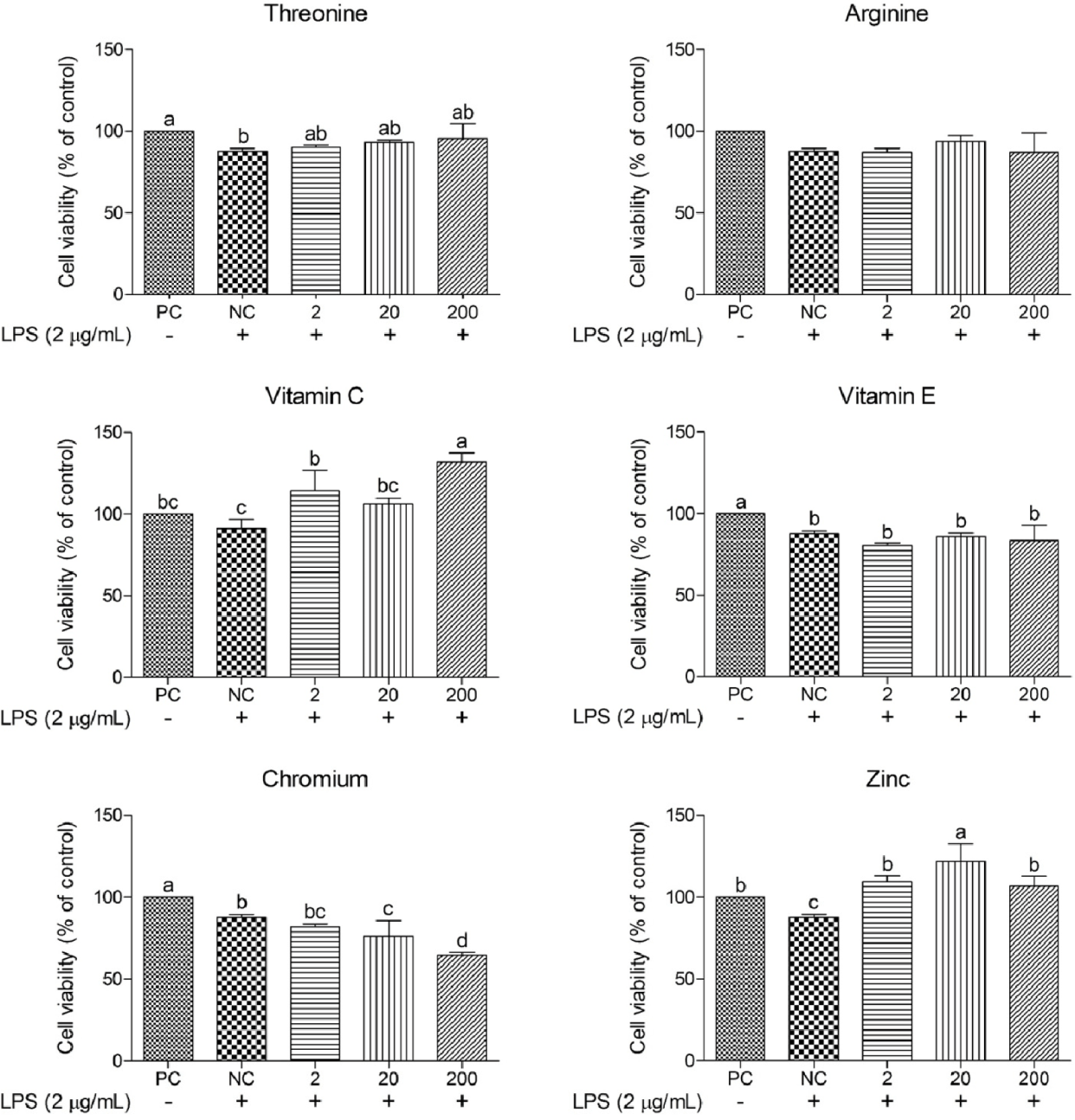
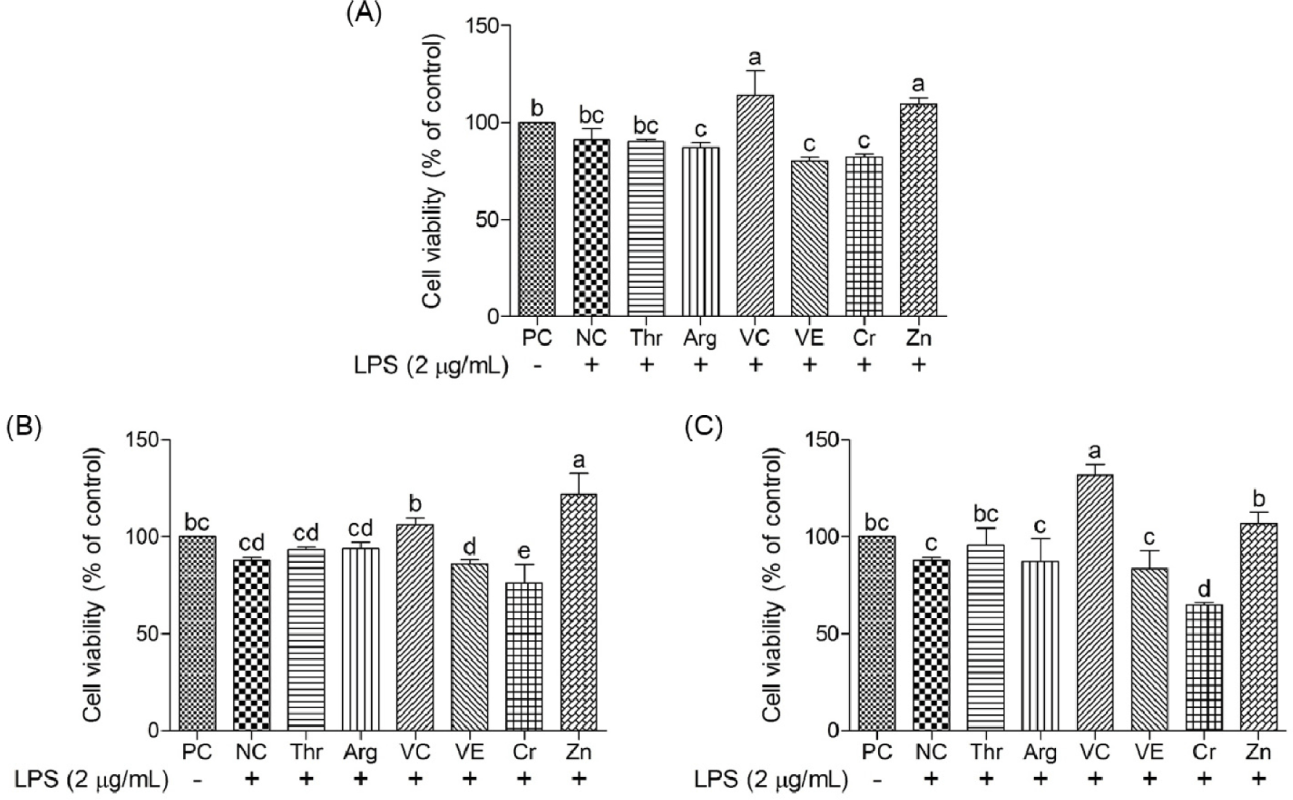
The cell viability of cIECs treated with different concentrations of LPS (0.25–2 μg/mL) was determined to confirm the optimal concentration for induced oxidative stress (Fig. 1). Cell viability after LPS treatment at 2 μg/mL was significantly less (p < 0.05) than that at other LPS concentrations; hence, this LPS concentration was selected for further study. Various functional nutrients, including Thr, Arg, vitamin C, vitamin E, Cr, and Zn, were evaluated for their potential to mitigate oxidative stress in cIECs. These functional nutrients were tested at concentrations of 2, 20, and 200 μg/mL for their impact on oxidative stress-induced cell viability (Figs. 2 and 3). Cell viability in the LPS-treated negative control (NC) was significantly less (p < 0.05) than that in the PC. However, Arg, vitamin E, and Cr did not exert significant effects on cell viability. Thr demonstrated a trend of cell-viability recovery to levels comparable to those of the PC; however, it did not exert a dose-dependent effect on cell viability. Vitamin C had significantly greater (p < 0.05) cell viability compared with the NC; in particular, at 200 μg/mL, it elicited greater (p < 0.05) cell viability than the PC. This suggests that vitamin C provides superior protection against oxidative stress. In addition, Zn also significantly increased (p < 0.05) cell viability at all concentrations compared with the NC; specifically, at 20 μg/mL, it exerted a greater (p < 0.05) effect than the PC. A concentration-dependent comparison of cell viability among treatment groups is presented in Fig. 3. Specifically, at 2 μg/mL, vitamin C exhibited a cell viability of 114.30 ± 10.18%; at 20 μg/mL, Zn achieved 122.00 ± 8.59% cell viability; and at 200 μg/mL, vitamin C demonstrated the greatest cell viability of 131.69 ± 4.66%.
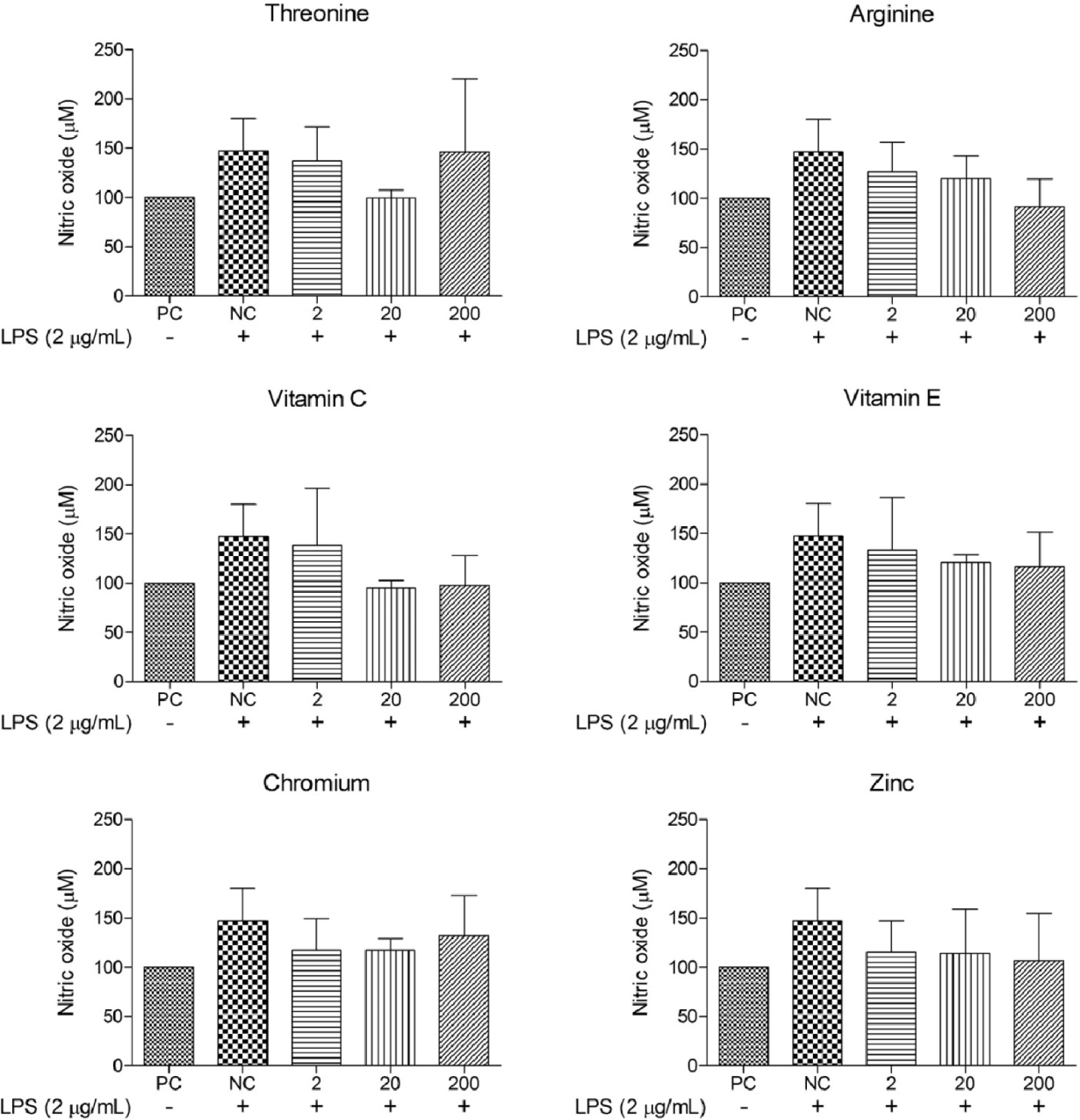
The effects of diverse functional nutrients and their concentrations on NO levels in oxidative stress-induced cIECs were evaluated (Figs. 4 and 5). The NO level in the NC was determined to be 147.24 ± 32.88 μM, approximately 47% greater than that in the PC.
This study evaluated the effects of different functional nutrients and their concentrations on HSP70 levels in oxidative stress-induced cIECs (Figs. 6 and 7). The HSP70 relative level in the NC group was 1.10 ± 0.16 ng/mL, approximately 10% greater than that in the PC group, indicating oxidative stress-induced increase. Certain functional nutrients yielded less (p < 0.05) HSP70 relative levels than the NC group, suggesting their effectiveness in mitigating oxidative stress. All functional nutrients, except Thr and vitamin C, significantly reduced (p < 0.05) HSP70 relative levels compared with the NC. Vitamin E, Cr, and Zn treatments exhibited particularly notable reductions, yielding significantly less (p < 0.05) HSP70 relative levels than those in the PC group, thereby indicating their superior potential for HSP70 suppression. However, vitamin C and Thr did not display significant changes in HSP70 relative levels compared with the NC. A concentration-dependent comparative analysis of HSP70 relative levels among treatment groups revealed that Arg, Cr, and Zn at 2 μg/mL elicited significantly decreased (p < 0.05) levels than the NC, while Cr and Zn at 20 μg/mL as well as vitamin E and Zn at 200 μg/mL followed suit (Fig. 7).
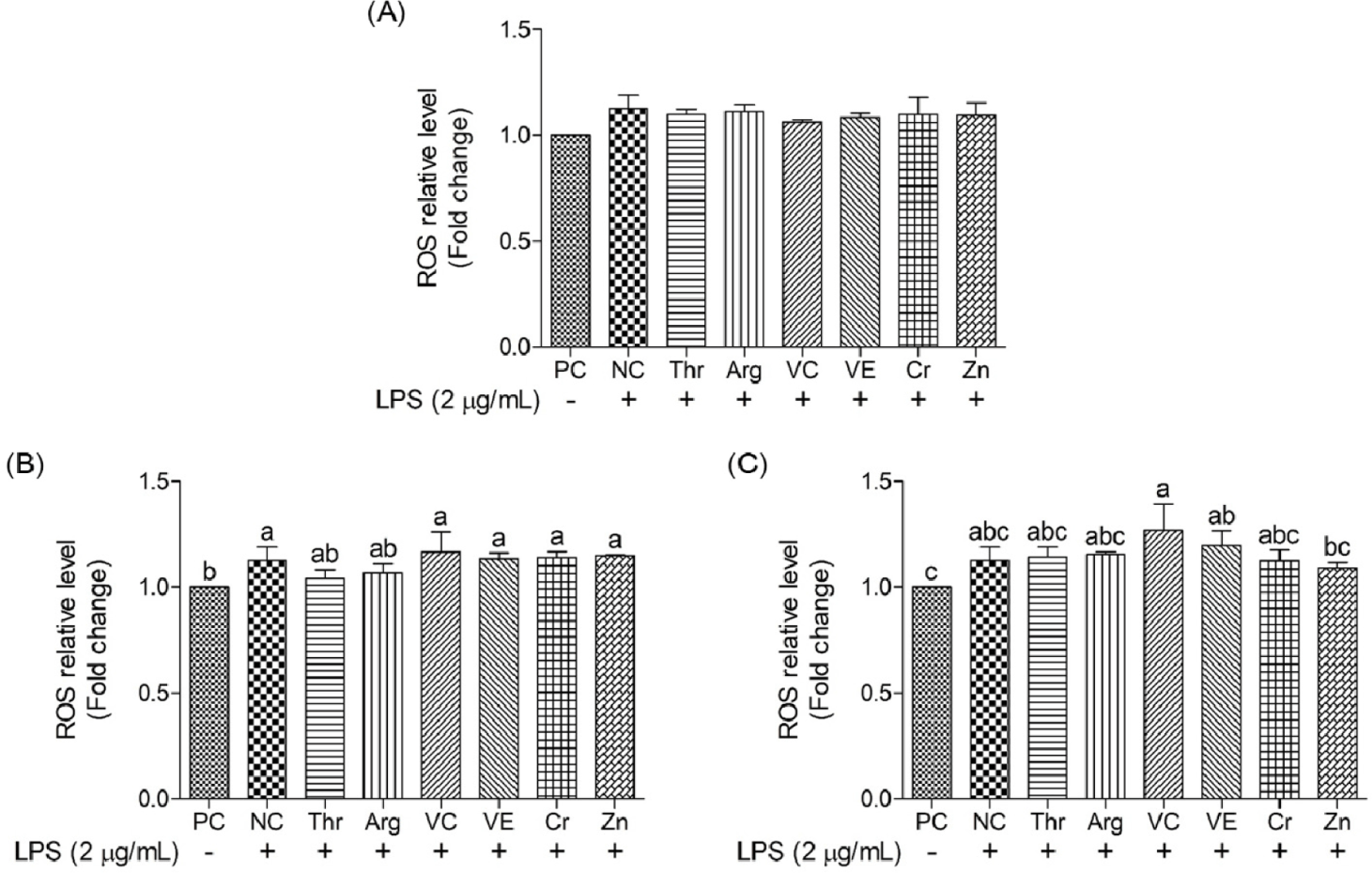
This study assessed the impact of functional nutrients on intracellular ROS levels in oxidative stress-induced cIECs (Figs. 8 and 9). In the NC group, the ROS relative level was 1.12 ± 0.06, a figure 12% greater than that in the PC group, indicating a significant increase (p < 0.05) in ROS owing to oxidative stress. However, these functional nutrients were no significant difference in ROS relative levels.
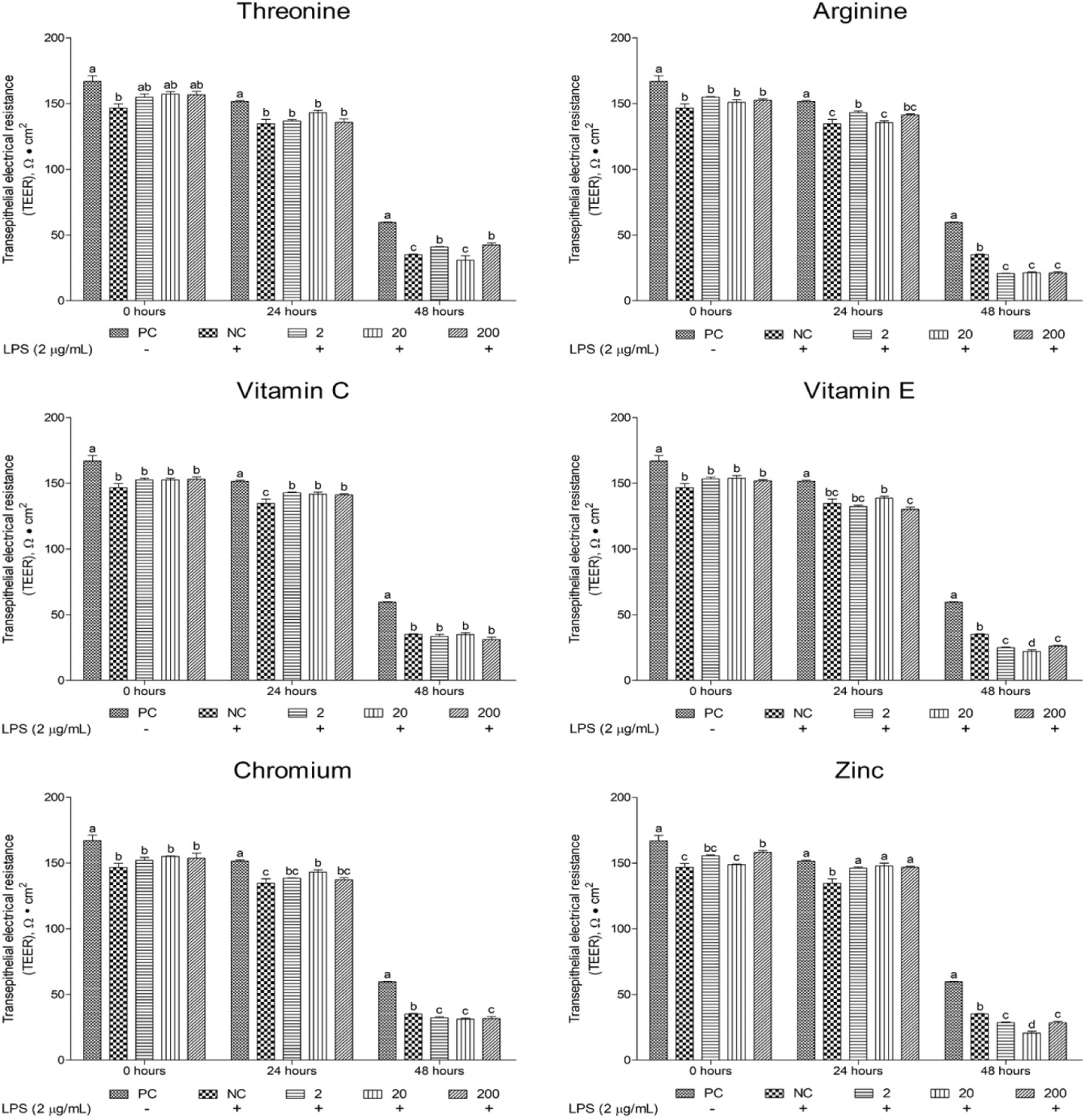
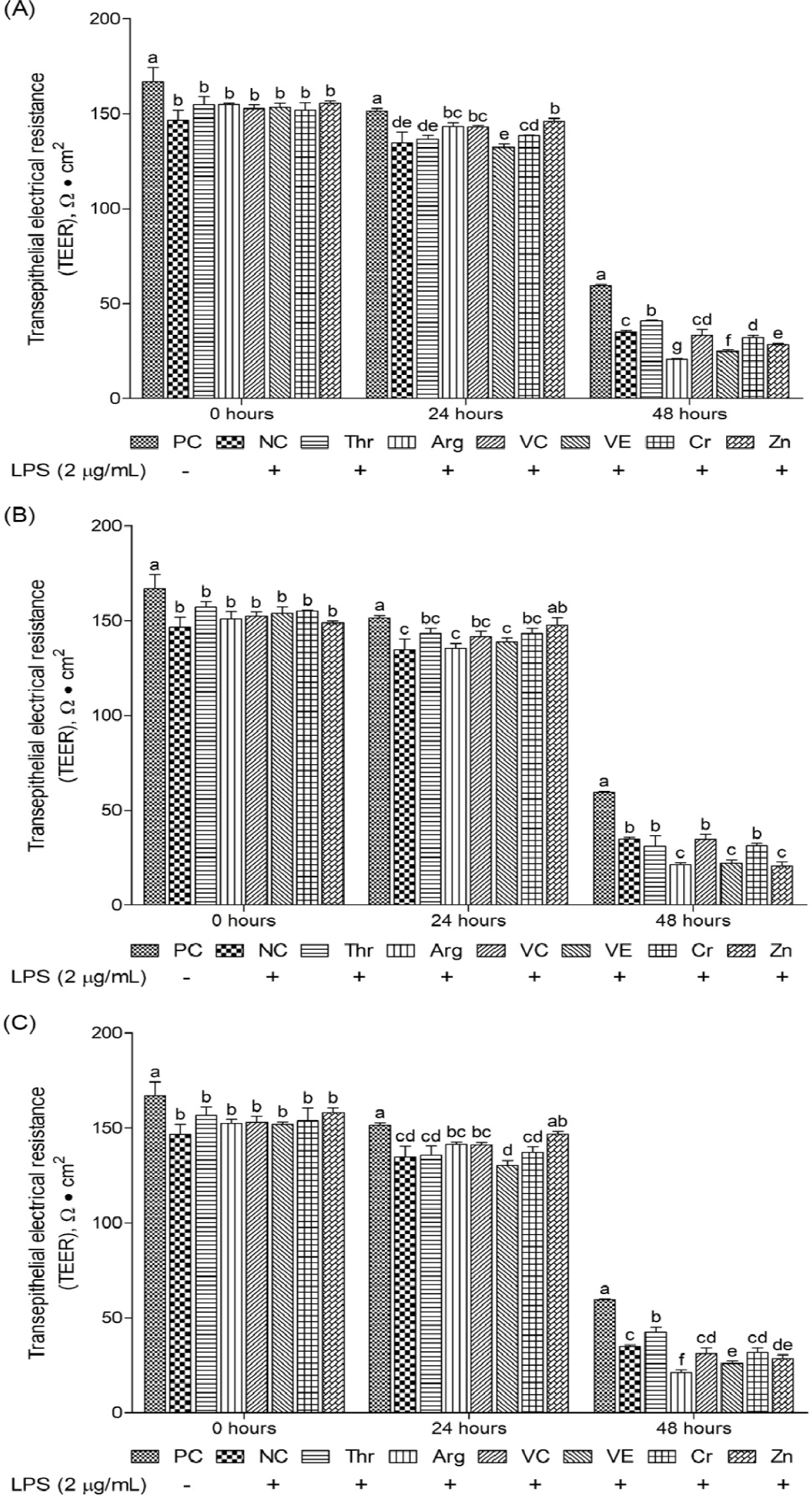
This study evaluated the effects of different functional nutrients and their concentrations on TEER recovery in oxidative stress-induced cIECs, focusing on tissue homeostasis and epithelial barrier robustness (Figs. 10 and 11). The initial (0 hours) TEER value for the NC group was 146.73 ± 5.14. At 24 and 48 hours post-treatment, the NC group’s TEER value significantly declined (p < 0.05) to 134.63 ± 5.749 and 35.1 ± 0.624, representing 7% and 77% reductions, respectively, compared with that at 0 hours. These results highlight the detrimental effects of LPS-induced oxidative stress on the epithelial barrier. Among the investigated functional nutrients, vitamin C and Zn significantly increased (p < 0.05) TEER levels at 24 hours compared with the NC. In addition, certain function materials increased (p < 0.05) TEER levels compared with the NC. In particular, Zn (24 hours) at all supplemental levels and Thr (48 hours) at 2 and 200 μg/mL were found to enhance (p < 0.05) TEER levels (Fig. 11).
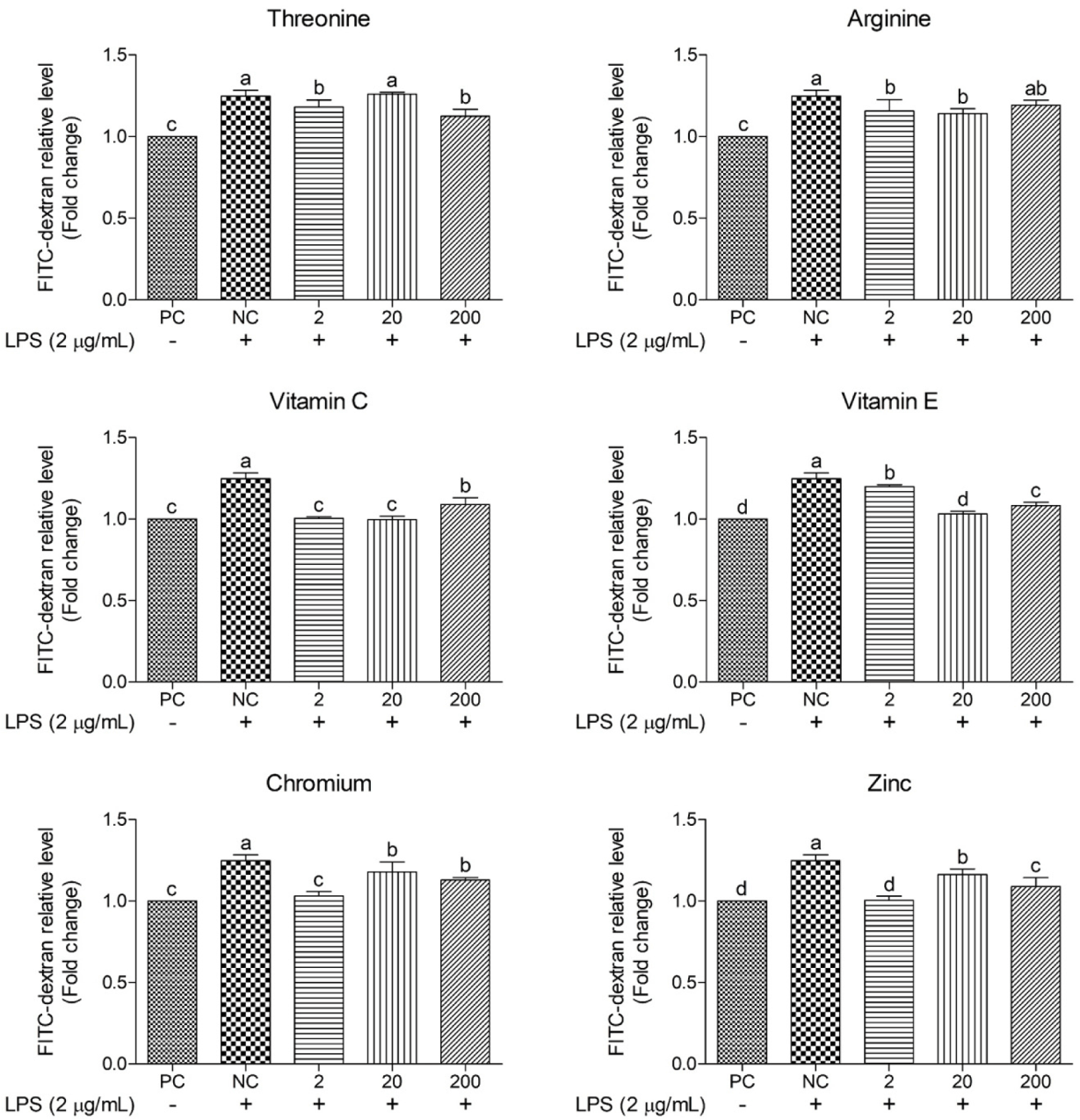
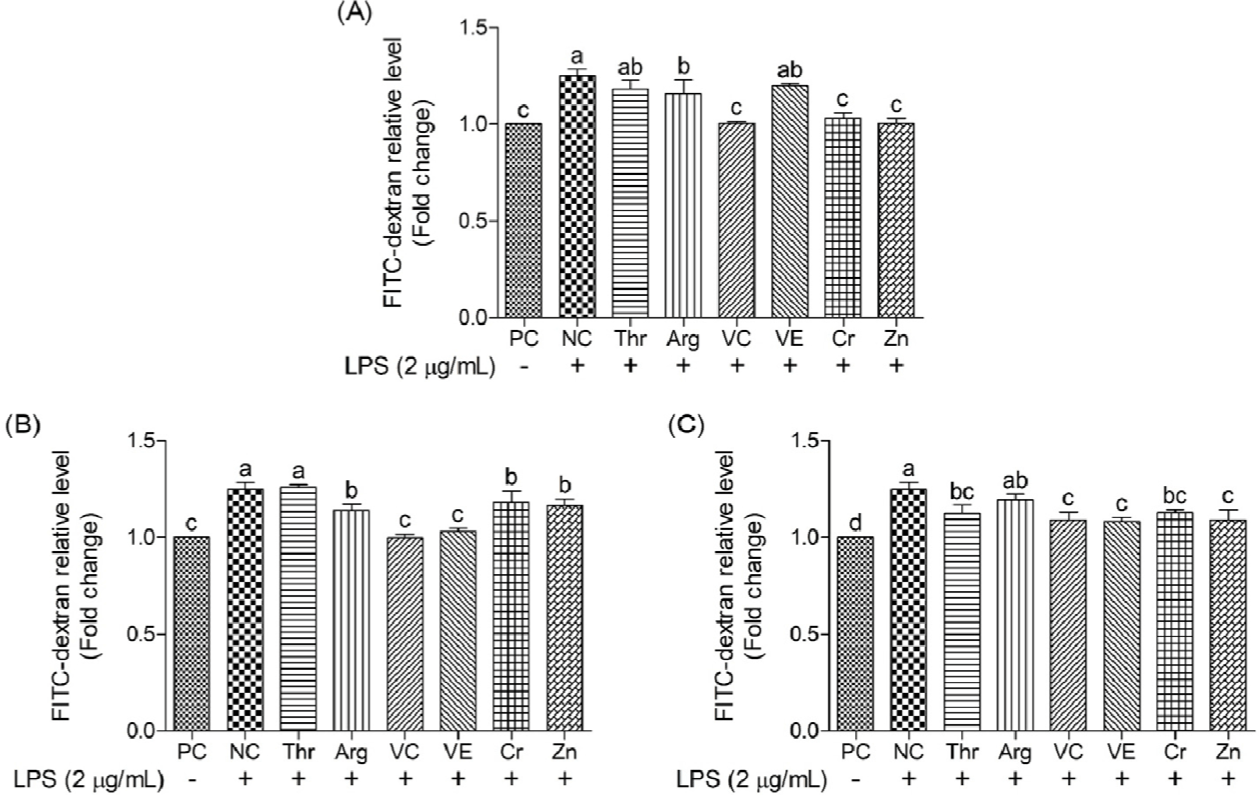
This study employed FITC–dextran to determine the protective effects of functional nutrients and their concentrations on oxidative stress-induced epithelial barrier damage (Figs. 12 and 13). The NC yielded 25% greater FITC–dextran levels than the PC, indicating significant barrier damage. On comparing treatment groups, vitamin C, vitamin E, Cr, and Zn consistently reduced (p < 0.05) FITC–dextran permeability across all concentrations, with certain concentrations exhibiting permeability levels similar to those of the PC. Thr (2 and 200 μg/mL) and Arg (2 and 20 μg/mL) resulted in significant reductions (p < 0.05) in FITC–dextran relative levels. Moreover, across all concentrations, most functional nutrients effectively reduced FITC–dextran permeability, with vitamin C and Zn demonstrating the greatest efficacy.
DISCUSSION
This study measured the effects of functional nutrients on cell viability among the functional nutrients, and the supplementation of vitamin C at 200 μg/mL and Zn at 20 μg/mL significantly enhanced cell viability. Vitamin C is a potent antioxidant and an excellent electron donor, effectively neutralizing ROS and free radicals [30]. Vitamin C treatment reportedly enhanced the regenerative capacity and cell viability of oxidative stress-induced IPEC-J2 cells [31]. Similarly, Zn functions as an antioxidant by competing for binding sites with oxidizable metals, lipids, proteins, and DNA, thereby maintaining the stability of sulfhydryl groups and promoting the synthesis of Cu–Zn SOD, a crucial antioxidant enzyme, to protect cells from oxidative damage [32,33]. Shao et al. [34] found Zn to upregulate the expression of zonula occludens-1 (ZO-1) protein by activating the PI3K/AKT/mTOR signaling pathway, thus enhancing intestinal epithelial barrier function and significantly increasing cell viability. These findings suggest that both vitamin C and Zn are particularly effective in improving oxidative stress-induced cell viability, with potential applications as protective functional nutrients. Further studies are warranted to elucidate their mechanisms of action and potential synergistic effects.
NO, a key marker produced during cellular inflammatory responses, is known to proliferate under oxidative stress conditions [35]. Vitamin C is a powerful antioxidant, and much research is being conducted on its inhibitory effects on NO production. Akolkar et al. [36] found vitamin C to effectively reduce nitric oxide synthase levels that had been elevated by doxorubicin-induced oxidative stress. Furthermore, vitamin C enhances the stability of the dimeric form of endothelial nitric oxide synthase, thereby reducing NO production. Based on these findings, vitamin C holds promise as a functional nutrient capable of suppressing NO level increases. This property suggests its potential application in mitigating inflammation and oxidative damage. However, in this experiment, vitamin C did not significantly reduce NO relative levels. Therefore, further studies are required to compare NO reduction effects of vitamin C at varying concentrations in cIECs under oxidative stress.
HSP70 is a representative stress protein whose expression increases in response to multiple stress types, including heat shock [37]. Inducible HSP70 is closely associated with the stress-resistance capacity of livestock, and serum HSP70 levels in animals, as a biomarker of heat resistance, can help determine the degree of cellular heat stress [38]. In this study, the supplementation of Arg, Cr, and Zn at 2 µg/mL in oxidative stress-induced cIEC reduced HSP70 relative levels. Arg plays a key role in supporting intestinal immunity and mucosal repair while also regulating HSP expression to maintain proper protein folding and function [39]. In addition, in fetal kidney cells exposed to Adriamycin™ (doxorubicin), Arg was associated with decreased HSP70 expression, suggesting its possible inhibition of the stress response under certain pathological conditions [40]. Additionally, in renal tubular cells, Arg has been shown to induce cellular stress, potentially altering HSP70 levels [41]. In this study, Arg downregulated HSP70 expression in oxidative stress-induced cIECs. However, its dose-dependent increase in HSP70 levels at greater concentrations indicates that it modulates HSP expression to maintain cellular homeostasis under stress. Exposure to Zn can inhibit HSP70 induction by affecting heat shock transcription factor activity, which correlates with reduced HSP70 levels in thermotolerant cells [42]. Li et al. [43] demonstrated that Zn supplementation, either in inorganic Zn or organic Zn forms, decreased HSP70 and heat shock protein 90 mRNA expression, thereby lowering HSP levels in hepatocytes. Therefore, Zn appears to modulate HSP expression in cIECs, thereby reducing HSP70 levels and consequently protecting cells from oxidative stress. In contrast, Cr has been reported to significantly increase HSP70 activity in Institute of Cancer Research mice [44]. The conflicting results may emanate from the attenuated cell viability observed in this study, implying that the significant decrease in HSP70 levels may not solely result from the direct Cr-induced downregulation of HSP expression levels but could also be influenced by reduced cellular activity. These results suggest that Arg, Cr, and Zn potentially provide a cost-effective means of reducing HSP70 levels.
ROS, such as hydroxyl radicals and hydrogen peroxide, are highly reactive molecules containing oxygen atoms. These molecules are naturally produced during cellular metabolic processes and are tightly regulated. However, excessive ROS levels may induce oxidative stress, leading to cellular damage [45]. Vitamin C acts as a radical scavenger and reducing agent, directly neutralizing ROS and regenerating vitamin E from its oxidized form, thereby enhancing its antioxidant activity [46]. Vitamin E serves an indispensable role as a chain-breaking antioxidant, protecting cell membranes from lipid peroxidation and oxidative damage [46]. However, both vitamins used this study did not show to significant protective effects against oxidative-induced cellular injury. It reportedly improves cell viability and tight-junction integrity in intestinal epithelial cells under oxidative stress conditions [25]. In addition, both vitamins demonstrate significant protective effects against oxidative-induced apoptosis and cellular injury. Further, a concentration-dependent comparative analysis of ROS levels among treatment groups revealed no significant difference between the NC and functional nutrients, except for Zn (200 μg/mL), regardless of concentration. Previous studies have demonstrated that Zn increases the expression of metallothionein, which binds to redox-active metals, like Fe or Cu, thereby preventing Fenton reactions [47]. Zn also directly reacts with H₂O₂ or hydroxyl radicals, neutralizing ROS before they can interact with DNA and induce oxidative stress [47]. However, the functional nutrients used in this study did not exhibit a superior role in inhibiting ROS production. Therefore, future studies should consider performing high-concentration treatment with functional nutrients to investigate their inhibitory effect on ROS.
TEER is a widely used indicator of cell-layer permeability and barrier integrity, measured using electrodes to assess electrical resistance across epithelial layers [48]. Greater TEER values indicate tight junctions and a healthier epithelial barrier. In the present study, Zn (24 hours) at all supplemental levels and Thr (48 hours) at 2 and 200 µg/mL significantly elevated TEER levels in oxidative stress induced cIECs. Previous research has demonstrated that zinc oxide significantly increases TEER levels by upregulating the mRNA expression of the tight junction protein ZO-1 in piglets infected with enterotoxigenic Escherichia coli (ETEC-K88) [49]. Similarly, Arg supplementation in media has been shown to sustain elevated TEER levels in IPEC-J2 over extended periods [50], while vitamin E treatment has been revealed to significantly increase TEER levels within 0–40 hours in IPEC-J2 cells [51]. However, in the present study, a significant increase in TEER level was exclusively observed with Zn treatment, whereas Arg and vitamin E supplementation did not have a notable impact on TEER increase. Tight junctions comprise transmembrane proteins, including claudins, occludins, and junctional adhesion molecules, and cytoplasmic scaffold proteins, including zonula occludens proteins [52]. Among these, most claudin proteins possess potential phosphorylation sites for serine and/or Thr residues within their cytoplasmic C-terminal domains [53]. Thr potentially influences the expression of claudin proteins, which are critical for tight junction integrity. Therefore, this study’s finding wherein Thr enhanced TEER after long-term treatment possibly correlates with claudin production via phosphorylation activation of Thr residues. These findings suggest that Zn may play a crucial role in restoring barrier function in cIECs under oxidative stress within 24 hours. In addition, Thr was found to be an advantageous material for tight junction health during long-term treatment.
FITC–dextran is an extensively used marker used to evaluate intestinal barrier integrity, especially in assessing epithelial permeability. FITC–dextran molecules (4–6 kDa) are typically unable to penetrate intact intestinal barriers unless damage occurs owing to infections, stress responses, or inflammation. An increase in FITC–dextran permeability indicates greater barrier damage and elevated permeability [54]. In our study, the supplementation of vitamin C and Zn in oxidative stress induced cIECs decreased FITC–dextran and showed similar levels with PC. Previous studies have demonstrated that high doses of vitamin C increase the mRNA expression of the tight junction protein ZO-1 and reduce elevated intestinal permeability in inflammatory bowel disease through its interaction with claudin-2 [55]. Likewise, Zn reportedly enhances cell differentiation and ZO-1 expression in Caco-2 cells by activating the PI3K signaling pathways, including the AKT and mTOR pathways, thereby contributing to reduced FITC–dextran permeability [34]. Based on these findings and previous studies, supplementation with vitamin C and Zn evidently promotes the expression of tight junction proteins, such as ZO-1, aiding in the recovery of LPS-damaged epithelial barriers and effectively decreasing FITC–dextran permeability in cIECs.
CONCLUSION
This study investigated the protective effects of functional nutrients in terms of enhancing cIEC barrier function and alleviating oxidative stress in chickens. Vitamin C and Zn in oxidative stress induced cIECs enhance cIEC proliferation compared to other candidate materials. Arg and Zn in oxidative stress induced cIECs effectively decrease HSP70 levels. However, none of the materials substantially decreased NO or ROS levels. Vitamin C and Zn in oxidative stress exposed to cIECs increase TEER and decrease FITC–dextran leakage, indicating enhanced barrier integrity. Prolonged treatment with Thr was particularly beneficial for tight junction health. Overall, this study suggests that amino acids (Arg and Thr), vitamin C, and Zn potentially serve as effective functional nutrients that enhance intestinal barrier function and protect against stress in poultry. Future research should focus on the in vivo validation and elucidation of the underlying molecular mechanisms.