
INTRODUCTION
Heat stress induces a series of behavioural and metabolic changes to maintain homeothermy in cattle; one of these behavioural changes leads to lower dry matter intake (DMI) [1]. Traditionally, lower DMI has been thought to be the main reason for lower milk production, milk protein synthesis, and ultimately milk protein yield in heat-stressed dairy cows [1–3]. However, paired feeding-restriction tests in non-heat-stressed and heat-stressed cows has found that lower DMI during heat stress could only partially explain decreases in milk production [4–5]. Several molecular mechanisms have been hypothesised to explain the reduction in milk synthesis activity in bovine mammary gland cells [4,6,7]. One study has demonstrated that the effect of heat stress on inhibition of milk protein synthesis in bovine mammary epithelial cells (BMECs) may be related to decreases in phosphorylation of 4E binding protein 1 (4EBP1), and not through the mechanistic target of rapamycin (mTOR) pathway [8].
In most cells, heat-stress and hyperthermia have been shown to induce apoptosis [9,10]. Several studies on BMECs have shown that heat stress-induced upregulation of apoptosis-related genes such as heat shock proteins (HSP70, HSPA5, and HSP90B1) [11], cysteinyl aspartate specific proteinase-3 (Caspase-3) [7], and Bcl-2-associated X protein (BAX) [12], and subsequent apoptosis of BMECs, could be contributing to lower milk protein synthesis.
Since methionine (Met) is one of the most limiting essential amino acids for milk protein synthesis [13,14], dietary supplementation with Met can be an effective way to improve milk production in dairy cows [15–17]. Dietary supplementation with 2-hydroxy-4-(methylthio)-butanoic acid (HMB; a pelletable hydroxy analogue of Met) has been shown to improve milk protein yields and nitrogen utilization efficiency by improving amino acid balance in metabolizable proteins and reducing deamination of absorbed amino acids [15]. Another report has shown that dairy cow diets supplemented with Met could significantly improve milk production by improving DMI and liver function [18]. In the context of heat stress, rumen-protected Met could help maintain milk protein production without affecting DMI and milk yield [19]. In vitro experiments have shown that genes linked to protein synthesis regulation (such as CSN1S1, CSN1S2, CSN2, CSN3,JAK2, and mTOR) were upregulated in Met-supplemented BMECs [20]. In addition, supplementation with Met had protective effects on BMECs under heat stress as it reduced apoptosis in these cells [21,22] and upregulated proliferation [8]. However, although Met supplementation has been shown to exert positive effects on the survival, proliferation, and protein synthesis of BMECs under heat stress, the molecular mechanisms of these processes, especially those involved in improving milk protein synthesis, are not entirely clear.
Here, we have explored the effects of different concentrations of Met supplementation on the protein synthesis of the BMEC line, mammary alveolar cells-large T antigen (MAC-T) under heat stress.
MATERIALS AND METHODS
MAC-T cells were kindly provided by Professor Songjia Lai (College of Animal Science and Technology, Sichuan Agricultural University, China) and cultured in 75 cm2 cell flasks (Thermo Scientific, Waltham, MA, USA) in 2 mL of growth medium (minimum essential medium [MEM], Thermo Scientific) with Earle’s balanced salt solution (EBSS, Thermo Scientific) supplemented with 10% (v/v) foetal bovine serum (FBS, Thermo Scientific), 5 mg/L insulin, 1 mg/L hydrocortisone, 5 mg/L transferrin, 5 μM ascorbic acid, 5 mM sodium acetate, 100 IU/mL penicillin, and 100 IU/mL streptomycin, 1 mg/L progesterone, 0.04% lactalbumin, and 0.04% α-lactose; all reagents were purchased from Sigma Aldrich (St Louis, MO, USA) unless otherwise specified) at 37°C in a humidified incubator containing 5% CO2. The cells were sub-cultured repeatedly until a confluency of 85%–90% was achieved in order to produce enough cell numbers for the experiments. Once this was achieved, cells were transferred to 6-well plates (Thermo Scientific) at a density of 1.2-1.5×105 cells in each well. Following this, the cells were cultured in FBS-free growth medium at 37°C for 12 hours and allowed to reach 85%–90% confluency in the 6-well plates as per methods previously described [8,18]. The cell growth medium was then removed and cells were washed three times with phosphate-buffered saline (PBS, Thermo Scientific). The washed cells were divided into four groups and supplied with 2 mL custom-made high-glucose devoid of all essential amino acids [DMEM], custom-made by Gibco, Carlsbad, CA, USA) supplemented with 100 IU/mL penicillin G and 100 μg/mL streptomycin; each group was further treated with four different mixtures of amino acids (Table 1). The four treatment groups were: (1) CON – control medium with a normal amino acid profile (60 μg/mL of Met), (2) MET70 – medium containing 70 μg/mL of Met (normal amino acid profile plus 10 μg/mL Met), (3) MET80 – medium containing 80 μg/mL of Met (normal amino acid profile plus 20 μg/mL Met), and (4) MET90 – medium containing 90 μg/mL of Met (normal amino acid profile plus 30 μg/mL Met). The treatments differed only in the amount of additional Met supplementation they contained; all other amino acid ratios were maintained at the normal amino acid profile levels. The cells in different treatments (n=6 replicates per treatment) were incubated at 42°C for 6 h. The incubation conditions were based on the similar work of Collier et al. [23] in which the mRNA abundance of heat shock protein 70 (HSP70) was significantly stimulated between 1 and 2 hours, and reached the peak within 4 hours after BMEC was exposed to 42°C. After this, cells were scraped from the wells, collected using a cell lifter in 1 mL of TRIzol reagent (Invitrogen, South San Francisco, CA, USA), and stored at −80°C until they were used for RNA extraction.
1) CON, normal amino acid profile (60 μg/mL of Met in medium); MET70, normal amino acid profile plus 10 μg/mL Met (70 μg/mL of Met in medium); MET80, normal amino acid profile plus 20 μg/mL Met (80 μg/mL of Met in medium); MET90, normal amino acid profile plus 30 μg/mL Met (90 μg/mL of Met in medium).
Total RNA from cells was extracted and purified using the Steady Pure Universal RNA Extraction Kit (Accurate Biotechnology, Hunan, China) as per the manufacturer’s protocol. A NanoDrop ND-2000 UV-VIS spectrophotometer (Thermo Scientific) was used to measure RNA concentrations at an optical density of 260 nm (OD260); RNA purity was assessed through OD260/OD280 ratios. In addition, the integrities of the RNA samples were confirmed via denatured agarose gel (1.5%) electrophoresis and integrity numbers for each sample were evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). RNA samples with 28S / 18S rRNA ratios > 1.8 were used for RT-PCR (real-time polymerase chain reaction) analysis [24]. Intact RNA samples (with RNA integrity number ≥ 7) with 1.8<OD260 / OD280 < 2.0 were used to generate labelled targets [25]. In this experiment, the OD260 / OD280 ratio of all RNA samples was > 1.9. Samples with RNA integrity numbers > 7 were diluted in RNase-free water to obtain solution of RNA at 100 ng/μL; these were used to obtain cDNA via reverse transcription using the PrimeScript™ RT-PCR reagent Kit with gDNA Eraser (Takara, Tokyo, Japan) according to the manufacturer’s protocol. The cDNA was diluted in a 1 : 4 ratio with RNase-free water for further experiments.
Quantitative real-time PCRs (qRT-PCRs) were performed in an ABI 7500 real-time thermocycler (Applied Biosystems, Foster City, CA, USA) according to previously reported protocols [26,27]. Briefly, 10 μL PCR reactions were prepared by mixing 4 μL of diluted cDNA with 6 μL of a mixture composed of 5 μL 1× SYBR Green Master Mix (Quanta, Gaithersburg, MD, USA), 0.3 μL each of 10 μM forward and reverse primers, and 0.4 μL RNase-free water. The reactions were performed under the following program for two-step amplification: initial denaturation at 95°C for 5 min, followed by 40 cycles of a two-step reaction (95°C for 15 sec and 60°C for 30 sec). Melting curves (95°C for 15 sec, 65°C for 15 sec, and 95°C for 15 sec) with continuous fluorescence measurements were subsequently constructed for each PCR product. The specificity of each qRT-PCR was evaluated by analysing the melting curve of the PCR products [28]. Amplification efficiency was calculated based on the slope of the melting line using the equation: E = 10 (−1/slope) – 1; the ideal range of values for E lie between 0.95 and 1.05 [25].
Primer sequences were designed using the Primer ExpressTM Software (Applied Biosystems) and validated with NCBI (National Center for Biotechnology Information) Primer-BLAST. The detailed list of primer sequences has been provided in Table 2. The reference genes [20,29] used for normalization of target gene levels were glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH), ribosomal protein S9 gene (RPS9), and the ubiquitously expressed prefoldin like chaperon gene (UXT); these genes were chosen after assessment by the geNorm application [30] as the most stable internal control genes for these experiments. The relative mRNA levels of target genes were determined by the 2−ΔΔCt method [31]. PCR experiments for each biological sample were repeated 3 times in a 384-well RT-PCR plate (Applied Biosystems).
CSN1S1, αS1-casein; CSN2, β-casein; CSN3, κ-casein; HSP70, heat shock protein 70; SLC7A5, solute carrier family 7 member 5; SLC34A2, solute carrier family 34 member 2; SLC38A3, solute carrier family 38 member 3; SLC38A9, solute carrier family 38 member 9; SLC38A2, solute carrier family 38 member 2; BCL2, B-cell lymphoma-2; BAX, Bcl-2-associated X protein; UXT, ubiquitously expressed prefoldin like chaperone; RPS9, Ribosomal Protein S9; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Western blotting was performed according to the protocols described previously by Feng et al. [32]. Briefly, total cell proteins from each of the 6 replicates per treatment were extracted by washing the cells 3 times with PBS, and lysing them on ice in RIPA lysis and extraction buffer (Thermo Scientific) supplemented with protease inhibitor cocktail (Thermo Scientific, Waltham, MA, USA). The supernatants from the lysates were collected after centrifugation (12,000×g for 5 min at 4°C) and protein concentrations were determined by using a bovine serum albumin (BSA) Protein Assay Kit (Thermo Scientific). Protein samples were then denatured at 95°C for 5 min and subjected to sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS‐PAGE). Subsequently, the separated proteins were transferred onto a polyvinylidene fluoride (PVDF) membrane (Bio‐Rad Laboratories, Hercules, CA, USA) in a semidry transfer assembly (Bio‐Rad Laboratories). The PVDF membranes were washed in Tris-buffered saline containing Tween 20 (TBST), and incubated at room temperature for 1 h in a blocking buffer solution of 1% BSA (Sigma Aldrich) in TBST with constant agitation. Following this, the membranes were washed with TBST and further incubated overnight at 4°C in a TBST solution with 5% non-fat milk and a primary antibody (listed in Table 3). After another round of washing with TBST, the membranes were incubated (with constant agitation) at room temperature for 1 h with a secondary antibody (HRP-conjugated goat anti-rabbit IgG, Abcam, Cambridge, UK). Finally, the PVDF membranes were treated with Clarity Western ECL Substrate (Bio‐Rad Laboratories) and bands were visualised using a ChemiDoc MP System (Bio‐Rad Laboratories). Band intensities were quantified with ImageJ software (National Institutes of Health, Bethesda, MD, USA), and the degree of phosphorylation in each band was calculated by the ratio of phosphorylated protein band intensity to total protein band intensity. The band intensity of the β-actin band was used as an internal control.
p-AKT, phosphorylated AKT; BCL2, B-cell lymphoma-2; BAX, Bcl-2-associated X protein; mTOR, mechanistic target of rapamycin; p-mTOR, phosphorylated mechanistic target of rapamycin kinase; RPS6K1, ribo-somal protein S6 kinase 1; p-RPS6K1, phosphorylated ribo-somal protein S6 kinase 1; RPS6, ribosomal protein S6; p-RPS6, phosphorylated ribosomal protein S6; HSP70, heat shock protein 70.
All statistical tests were performed using SAS Software version 9.2 (SAS Institute, Cary, NC, USA). The Shapiro–Wilk test was also used to evaluate if all variables were normally distributed. One-way analysis of variance (ANOVA) tests were used to evaluate the differences between the mean values of normally distributed data; Further post-hoc analysis was performed by comparing the means with each other using the Student-Newman-Keul’s multiple comparison test. The mean values of mRNA and protein levels in the control group (CON) was set to 1.00. For all statistical tests, p-value < 0.05 was considered to be significant.
RESULTS
As compared to the CON, the mRNA levels of B-cell lymphoma-2 (BCL2) were significantly higher in both MET70 and MET80 (p < 0.05 for each; Fig. 1), whereas, the mRNA levels of BAX were significantly lower in both MET80 and MET90 (p < 0.05). In addition, mRNA levels of cysteinyl aspartate specific proteinase-9 (Caspase-9) were also observed to be significantly lower than those of the CON in in MET80 (p < 0.05). Additional Met supplementation did not seem to affect the mRNA levels of HSP70 as these levels were similar in the CON, MET70, MET80, and MET90.
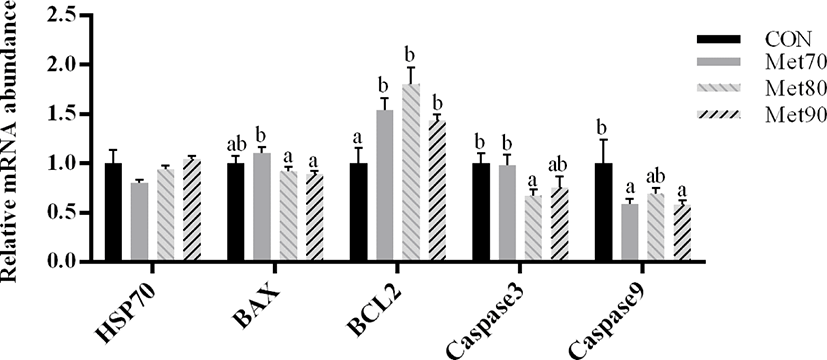
Compared to the CON, the protein levels of HSP70 were significantly higher in MET90 (p < 0.05; Fig. 2); similarly, levels of BCL2 were significantly higher in MET80 (p < 0.05) as compared to the CON. The protein levels of BAX were significantly lower in MET80 and MET90 (p < 0.05) than those in CON and MET70.
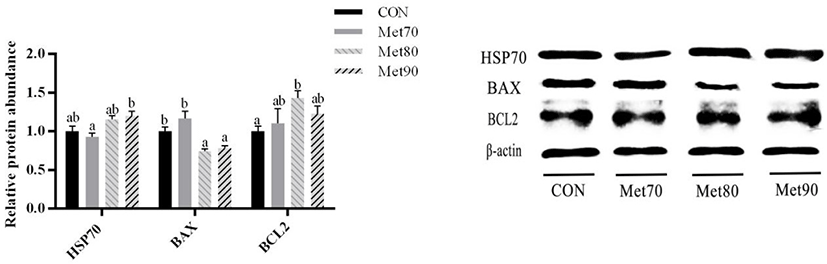
The protein levels of total mTOR (t-mTOR), total ribosomal protein S6 (t-RPS6) and serine-threonine protein kinase (t-AKT) were significantly higher in MET80 and MET90 as compared to those in the CON and MET70 (p < 0.05 for each comparison; Fig. 3A). Compared to its levels in MET90, the levels of RPS6 proteins were significantly higher in MET80 (p < 0.05). The levels of ribosomal protein S6 kinase B1 (t-RPS6KB1) proteins were significantly lower in MET70 as compared to those in the CON (p < 0.05). The levels of phosphorylated mTOR (p-mTOR), phosphorylated ribosomal protein S6 (p-RPS6), and phosphorylated serine-threonine protein kinase (p-AKT) proteins were significantly higher in MET80 and MET90 as compared to those in the CON (p < 0.05 for each; Fig. 3B). The levels of phosphorylated ribosomal protein S6 kinase B1 (p-RPS6KB1) proteins were significantly higher in MET80 than those in the CON and the other treatment groups (p < 0.05 for each comparison). As compared to the CON, the ratios of phosphorylated to total mTOR and ratios of phosphorylated to total RPS6 proteins were higher in MET80 (p < 0.05 for each; Fig. 3C). Similarly, the ratios of phosphorylated to total AKT proteins were higher in MET80 and MET90 (p < 0.05 for each) as compared to those in the CON. The ratios of phosphorylated to total RPS6KB1 proteins were higher than those in the CON for all additional Met supplementation treatments (p < 0.05).
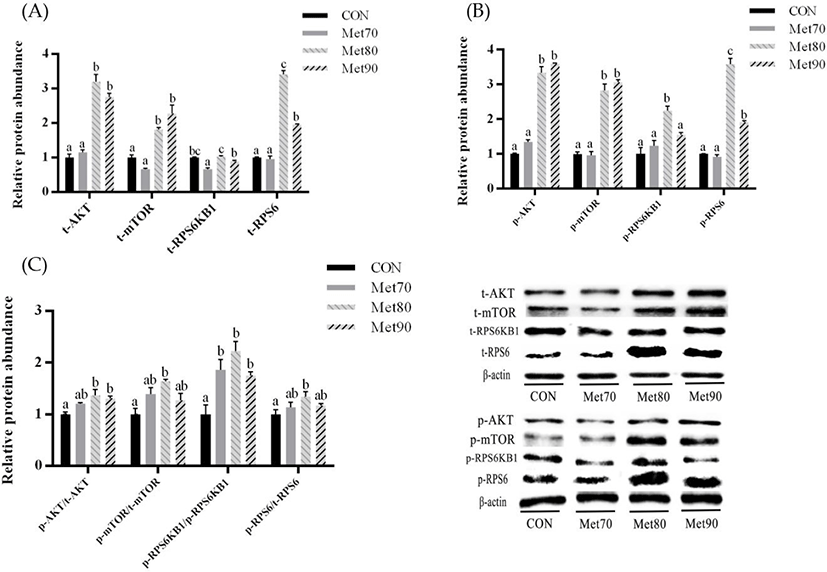
Supplementation with Met led to increases in the mRNA levels of some casein and amino acid transporters (AATs) in MAC-T cells under hyperthermic conditions, as shown in Fig. 4. Compared to the CON, the αS1-casein (CSN1S1), β-casein (CSN2), and solute carrier family 38 member 2 (SLC38A2) mRNA lever was significantly higher in MET80 (p < 0.05 for each); however, levels of solute carrier family 7 member 5 (SLC7A5) mRNA were significantly lower (p < 0.05). Compared to the CON, the SLC38A2 mRNA levels for MET90 were significantly higher (p < 0.05), whereas those of SLC7A5 mRNA were significantly lower (p < 0.05). In addition to these results, solute carrier family 38 member 9 (SLC38A9) mRNA levels in MET70 and SLC38A3 mRNA levels in MET80 were significantly higher than those of the CON (p < 0.05 for each). Additional Met supplementation did not affect the mRNA levels of κ-casein (CSN3) and solute carrier family 34 member 2 (SLC34A2) as these levels were similar in the CON, MET70, MET80, and MET90.
DISCUSSION
In mammalian cells, hyperthermia produces a series of negative effects on cellular functions—including inhibition of DNA synthesis, inhibition of RNA transcription, denaturation of proteins, and misaggregation of proteins—all of which can severely inhibit normal cellular activity and proliferation [33]. The durations and intensities of these negative effects determine the subsequent fate of the cells (adaptation and survival or apoptosis) under hyperthermia [34]. The inhibition of milk protein synthesis under heat stress is likely to be caused due to reductions in the numbers and activities of mammary cells in heat-stressed cows [5].
Apoptosis, which occurs as programmed cell death controlled by gene expression [22], requires the mobilization of a variety of pro- and anti-apoptotic molecules, including HSPs [35], Bcl-2 family members [36], caspases [37], etc. The accumulation of protein aggregations in cells is a key trigger for apoptosis. As protein aggregation occurs, there is an increase in the expression of HSPs as the cell attempts to remove the aggregated proteins to prevent nuclear deformation and cell death, etc. [38]. HSP70 is a highly inducible cytoprotective protein frequently used as a biomarker for cellular stress. An increase in HSP70 expression is a typical response to heat shock as the heat-stressed cell tries to maintain cellular metabolic activity and repair itself [39]; in addition, HSP70 can regulate apoptosis by regulating the expression of apoptosis-related genes [40].
One study has reported that in primary BMECs incubated at 41°C for 24 h, the expression of HSP70 increased significantly during the first 1 to 2 h of hyperthermia, then fell to basal levels after 8 h; during this time, genes involved in cell structure, metabolism, biosynthesis, and intracellular transport maintenance were downregulated [23]. The study demonstrated that after 8 h at 41°C, BMECs appeared to lose thermotolerance and that the cells began to undergo apoptosis. In another study on BMECs, mRNA levels of BAX were found to have doubled under hear stress; the same study also found that the heat stress caused the mRNA ratios of BAX/BCL2 to increase by 2.19 times [12]. These reports indicate that the changes in the expression profiles of pro- and anti-apoptotic factors were likely due to adaptive responses that BMECs resort to when exposed to hyperthermia.
The positive effects of Met supplementation on HSP70 gene expression in cells or tissues under heat stress, oxidative stress, and other stress conditions have been previously reported [22]. Under hyperthermia, an increase in the supply of amino acids (including Met) to BMECs could contribute to the synthesis of cytoprotective proteins such as HSP70 [8]. In this study, although we found that supplementation with Met had no effect on the mRNA levels of HSP70 in MAC-T cells under hyperthermic stress, we found that HSP70 protein levels were higher with Met supplementation (CON vs MET90 and MET80; Fig. 2). This may be explained by the fact that additional Met supplementation was beneficial to the accumulation of HSP70 in MAC-T cells during hyperthermia for 6 hours, while did not affect the further transcription of HSP70. Hence, it can be inferred that additional Met supplementation stimulated HSP70 expression only occured under short-term hyperthermia.
Supplementation with Met has been shown to inhibit BMEC apoptosis under heat stress; this was seen to be accompanied by increases in the expression of BCL2, while the expression levels of BAX and Caspase-3 were found to be decreased [20]. Met supplementation was also found to decrease the BAX/BCL2 ratios in BMECs under heat stress [22]. We have obtained similar results in this study; we find that supplementation with Met lowered both mRNA and protein levels of Caspase-9. We also found that additional Met supplementation upregulated BCL2 expression, whereas, the expression of Caspase-3 was downregulated. These results suggest that additional Met supplementation could deter the onset of apoptosis induced by hyperthermia in MAC-T cells; this, in turn, may contribute to an improvement in milk protein synthesis in heat-stressed MAC-T cells.
In nonruminants, various amino acids have been proved to be activators/regulators of the mTOR signalling pathway. Specific essential amino acids (eg., leucine, arginine, and Met) are not only important substrates in milk protein synthesis, but are also regulatory factors for various cellular functions as they can activate or inhibit various signalling pathways [41]. Many reports have confirmed that the mTOR signalling pathway is a crucial nutrition-sensitive pathway in amino acid-regulated mammary cell proliferation and casein synthesis [42–46]. As a pivotal upstream regulator of the mTOR signalling pathway [47], AKT regulates mTOR activity by sensing signals related to cell differentiation, proliferation, and apoptosis [48]. Phosphorylated AKT activates the mTORC1 complex by inhibiting the tuberous sclerosis (TSC) complex or by directly phosphorylating mTOR [49]. Without heat stress, Met supplementation in primary BMECs caused no changes to the levels of total and phosphorylated AKT [46]. However, under heat stress, Met supplementation raised levels of mTOR and AKT which had been suppressed by heat stress [8]. In this study, we found that in BMECs under heat stress, the levels of total and phosphorylated proteins, as well as the phosphorylation ratios of AKT were higher in the groups supplemented with Met (MET80 and MET90).
As a well-known nutrition-sensing signalling pathway related to protein synthesis, the activity of the mTOR pathway was found to be lowered in response to heat stress [42,50]. In accordance with different intracellular signals, mTOR binds to several proteins to form two distinct multi-protein complexes termed as mTORC1 and mTORC2. The mTORC1 complex is able to sense various signalling molecules induced by the presence/absence of nutrients and growth factors, as well as the cell’s energy status, to regulate cellular proliferation, growth, angiogenesis, and autophagy by controlling protein synthesis [51-52]. The mTORC1 complex regulates protein synthesis via phosphorylation to inactivate eukaryotic translation initiation factor 4E binding protein 1 (EIF4EBP1, a repressor of mRNA translation) and activate RPS6KB1 [53]. RPS6 is a component of the 40S ribosomal complex required for the initiation of mRNA translation [53], which is regulated by mTORC1/ RPS6KB1, and regulates the process of initiation during mRNA translation and protein synthesis [54]. In previous reports, the gene [55] and protein [11] expression levels of mTOR were found to be downregulated in heat-stressed BMECs. In addition, the phosphorylation levels of RPS6KB1 and eukaryotic elongation factor 2 (eEF2, a downstream factor of mTOR, whose levels are inversely related to those of mTOR) were found to be higher and lower, respectively, during hyperthermic stress [42]. Salama et al. [8] observed that heat stress had no effect on the phosphorylation of RPS6KB1 and RPS6, although it reduced mTOR protein levels and increased EIF4EBP1 mRNA levels. These studies indicate that the mTOR signalling pathway is inhibited in BMECs under heat stress, and probably contributes directly to the reduction in milk protein synthesis induced by hyperthermia.
In our study, Methionine supplementation (as seen in MET80) led to higher levels of phosphorylation in mTOR and proteins downstream of the mTOR pathway (including RPS6KB1 and RPS6) in BMECs under heat stress. These results indicated that additional Met supplementation could activate the mTOR signalling pathway in MAC-T cells under hyperthermia, which may rescue milk protein synthesis under heat stress. Furthermore, our results show that additional Met supplementation at a concentration of 20 μg/mL (80 μg/mL of Met in medium) could be optimal to reduce the deleterious effects of heat stress.
AATs on the cell surface play crucial roles in the transmission of information about nutrient availability in the external environment to the cell interior. These transporters can modulate the signal transduction occurring downstream of amino acid receptors by regulating intracellular amino acid concentrations [56]. For instance, the proteins from the solute‐linked carrier 36 (SLC36) family [57] and SLC38A9 [58] have been proved to be amino acid sensors that regulated mTORC1 signalling after they receive signals from binding to amino acids. The transcriptional control of AATs in mammary gland cells is directly and indirectly modulated by the lactation stage in dairy cows [59] and amino acid composition in metabolic proteins [20]. In addition, it was also found that during heat stress, there were increased rates of synthesis of structural proteins, pro-, and antiapoptotic proteins involved in restoring normal cellular function and directing cellular remodelling [60]; this increase automatically increased the availability of amino acids and AATs to meet the requirements for producing more cytoprotective proteins (e.g., HSPs).
In our study, we detected an increase in the levels of SLC38A2 and SLC38A3 mRNAs in heat stressed cells supplemented with Met (MET80 and MET90 groups), which is an expected and logical outcome given our previous results that showed upregulation of mTOR phosphorylation. The two proteins, SLC38A2 and SLC38A3, are AATs belonging to two subgroups of the SLC38 family, and have substrate specificities for glutamine, histidine, and aspartate [61]. In most cases, the activity levels of the mTOR pathway were consistent with the levels of SLC38A2 [62] and SLC38A3 [63]. However, in our study, we also found that additional Met supplementation did not affect the gene expression levels of SLC34A2, although the expression levels of SLC7A5 were downregulated. The gene SLC7A5 encodes AAT for the transmembrane exchange of neutral amino acids [64], and gene SLC34A2 encodes sodium-driven phosphate cotransporter for the transport of inorganic phosphate into cells [65]. Dong et al. [20] and Hu et al. [46] have found that Met supplementation could trigger a negative feedback effect due to which the increased availability of Met reduces the absorption of some amino acids. We observed a similar effect when we observed that the mRNA levels of SLC38A9 (an arginine sensor) first increased and then decreased with higher Met supplementation levels. Our results indicate that Met levels may regulate the entry of amino acids into the cells by affecting the expression levels of AATs (including SLC38A2, SLC38A9, and SLC7A5) [66], which in turn, modulate mTOR activity.
In milk, the four main isoforms of casein (including α-s1 casein, α-s2 casein, β-casein and к-casein) make up approximately 80% of the total protein content; the remaining milk protein components are composed of whey proteins. Since the MAC-T cell line does not produce milk components, the gene expression levels of casein were used as a proxy for measuring milk protein yield [67]. We have used the mRNA levels of casein as a proxy to measure the effects of additional Met supplementation on milk protein synthesis. Supplementation with Met has been shown to upregulate the expression of the casein genes (CSN1S1, CSN1S2, CSN2, CSN3) in BMECs [56]. In our experiments, we found that additional Met supplementation led to increased CSN1S1 and CSN2 mRNA levels in MAC-T cells exposed to hyperthermia.
Overall, our results indicate that Met levels control the entry of amino acids into cells by regulating the transcriptions of AATs. As substrates for protein synthesis, these amino acids could regulate the intracellular AA composition to get command of milk protein synthesis. Besides, these amino acids may act on the mTOR pathway to regulate intracellular amino acid compositions, and in turn, affect milk protein synthesis.
CONCLUSIONS
This study demonstrates that supplementation with Met may be used to improve milk protein synthesis under hyperthermia (42°C). Our results show that additional Met supplementation could inhibit apoptosis, activate the AKT-mTOR-RPS6KB1 signalling pathway, and regulate the gene expression of AATs; all these effects are likely to support the transcription of the casein genes which act as a proxy to measure milk protein synthesis. The best results were obtained with additional Met supplementation at a concentration of 20 μg/mL (80 μg/mL of Met in medium). Overall, these results indicate that the regulation of milk protein synthesis under hyperthermia likely depends on the levels of amino acids entering cells, which are in turn, regulated by Met levels. Our study provides evidence that Met supplementation in the feed of heat-stressed dairy cows may help in improving milk protein synthesis by affecting amino acid supply levels to mammary gland cells.