INTRODUCTION
The immune response is a system through which living organisms distinguish between external pathogens and normal cells, and the smooth operation of this system is essential for maintaining health. Recent research has increasingly interested on investigating the impact of microbiome treatments on the immune system, underscoring the significance of this field of study.
Typically, there are Lactobacillus strains and Bifidobacterium. Lactobacillus promotes the secretion of the anti-inflammatory cytokine interleukin-10 (IL-10) in the gut, thereby regulating IL-10-mediated immune responses and inhibiting excessive inflammation [1]. Furthermore, Lactobacillus inhibits the growth of harmful gut bacteria such as Clostridium difficile, while promoting the growth of beneficial bacteria like Bifidobacterium, thereby regulating gut microbiota balance and suppressing inflammatory responses. The Bifidobacterium, similar to Lactobacillus, promotes the secretion of anti-inflammatory cytokines (IL-10, Tumor necrosis factor beta [TGF-β]) and inhibits the production of pro-inflammatory cytokines (tumor necrosis factor alpha [TNF-α], Interleukin-6 [IL-6], and Interleukin 1 beta [IL-1β]) [2]. Additionally, Bifidobacterium enhances the gut barrier function, preventing the translocation of lipopolysaccharide (LPS), a toxic substance produced by harmful gut bacteria, into the bloodstream, thereby reducing inflammation [3]. Numerous clinical studies have demonstrated the efficacy of Bifidobacterium breve and Bifidobacterium longum in treating inflammatory diseases, including IBD.
Microbiome treatment is primarily achieved through probiotics [4], prebiotics [5,6], and even the transplantation of specific microbial groups [7]. Such interventions can activate beneficial microbial communities and regulate immune responses, potentially contributing to the prevention or treatment of various immune-related diseases.
Porphyromonas gingivalis is a Gram-negative bacterium known to be closely associated with periodontal diseases such as gingivitis and periodontitis [8]. Periodontitis typically begins when pathogens infect the space between the tooth and the gum known as the sulcus. Infections caused by P. gingivalis lead to inflammation in gum tissues, tissue destruction, and the promotion of osteoblast resorption in the alveolar bone [9]. The P. gingivalis penetrates into periodontal tissues and releases various outer membrane vesicles that can trigger innate immune responses, including inflammation [10,11]. LPS is one of the key factors among these outer membrane vesicles in the development of periodontitis [12]. The outer membrane in P. gingivalis LPS (PG-LPS) plays a crucial role in mediating inflammation and inducing cells to secrete pro-inflammatory cytokines [13]. For this reason, most of research utilize LPS to study microbiome infection and colonization.
The RAW 264.7 cell line is an immune cell line derived from murine macrophages, isolated from the Abelson murine leukemia virus (Abelson) and widely employed as a model cell line in inflammation and immune regulation research [14–16]. Research on LPS reduction using RAW 264.7 cells can provide valuable insights into the management and treatment of inflammatory conditions. For this reason, numerous previous studies have been conducted, including research on the anti-inflammatory effects using various compounds, especially natural product extracts, antioxidants, and anti-inflammatory drugs, following stimulation with LPS [17,18], studies identifying signaling pathways triggered by LPS, such as the TLR4-NF-κB pathway [19], modulation of immune responses, such as the activation of immune regulators (e.g., Treg cells, Th17 cells) and the expression of cell adhesion molecules [20,21], as well as investigations into the role of mitochondrial dysfunction and oxidative stress induced by LPS [22,23].
The aim of this study was conducted to discover a novel microbiome associated with the reduction of PG-LPS known to cause periodontitis and to identify the regulatory signaling pathways. In order to investigate the potential alleviating effects of periodontitis, we intend to utilize RAW 264.7 cells, commonly employed in immunological research, and treat them with PG-LPS. Subsequently, we plan to treat these cells with various microbiome strains derived from Canis familiaris Canine. The obtained result in this study will provide valuable foundational insights for future research into the mechanisms of canine periodontitis alleviation.
MATERIALS AND METHODS
Five healthy dog (average age: 4.5 years old) and five periodontitis infected dog (average age: 10.2 years old) were used in the study. Oral microbiome samples were collected by DNA/RNA Shield SafeCollect swab collection kit (Zymo Research). Swabs were placed into 2 mL of 0.1% buffered peptone water (BPW; Difco) medium as transport medium and oral microbiomes was immersed into 1 mL of de Man-Rogosa-Sharpe (MRS) broth to preferentially isolate lactobacilli. All swab samples were sent for laboratory analysis. Each swab was then immersed into 1 mL of MRS. All samples were stored at –80°C before molecular biological analysis.
The Mouse Macrophage RAW 264.7 cells were maintained and sub-passaged in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% antibiotic–antimycotic (Penicillin-Streptomycin [10,000 U/mL], Gibco). The cells were cultured at 37°C in a humidified atmosphere with 5% CO2. Routine medium changes were performed three times a week. Cells at 70% to 80% confluency were gently washed twice with PBS and harvested using 0.25% trypsin-EDTA (Gibco) for expansion. LPS from P. gingivalis were purchased from InvivoGen, and were used to induce inflammation response. To induce LPS stimulus, RAW 264.7 cells were incubated at 80% confluency and treated 1 µg/mL of LPS for 24 hours. To investigate the anti-inflammatory effects of the candidate microbiome, microbiome samples were treated by multiplicity of infection (MOI). The microbiome were treated simultaneously with LPS.
The Mouse Macrophage RAW 264.7 cells were seeded at 1 × 104 cells per well in 96-well plates and incubated for 24 hours. Then, the cells were treated with LPS from P. gingivalis various concentrations (125 ng/mL, 250 ng/mL, 500 ng/mL, and 1 μg/mL) for 24 hours. The level of nitric oxide (NO) produced in the culture medium was determined using Griess reagent (Promega). The Griess assay is one of the most common methods for quantifying NO. A Nitrite Standard reference curve prepared for accurate quantitation of NO levels in the Dulbecco’s Modified Eagle’s Medium (DMEM), which was supplemented with 10% fetal bovine serum and 1% antibiotic–antimycotic, and used for experimental samples. The 100 µL of culture supernatants was mixed with 50 µL of sulfanilamide solution and incubated for 8 minutes at room temperature, while being protected from light. And dispense 50 µL of N-1-napthylethylenediamine dihydrophloride (NED) solution to all wells. After a 20-min incubation period, protected from light, the absorbance at 540 nm was measured using a microplate reader (Tecan Spark).
RAW 264.7 cells were cultured in DMEM medium supplemented with 10% FBS and 1% penicillin-streptomycin. Cells were maintained at 37°C in a humidified incubator with 5% CO2. For each experiment, cells were seeded at a density of 3 × 10⁵ cells/well in 6-well plates and allowed to adhere for 24 hours. Following this incubation cells were treated with 1 µg/mL PG-LPS and candidate microbiome for an additional 24 hours to analysis apoptosis. Negative control wells were maintained without the inducing agent to assess staining specificity. After treatment, cells were washed twice with cold PBS to remove residual medium and inducers. Cells were then fixed with 4% paraformaldehyde for 15 minutes at room temperature. Following fixation, cells were washed with PBS and stained with DAPI to visualize nuclei. To distinguish apoptotic and necrotic cells, Annexin V-FITC and propidium iodide (PI) staining were performed. Annexin V conjugate and 100 µg/mL PI working solution were added to the cells, followed by incubation at room temperature for 15 minutes in the dark. Cells were subsequently washed with 1X annexin-binding buffer to remove excess staining reagents. Stained cells were observed under a fluorescence microscope equipped with appropriate filters to visualize DAPI, Annexin V, and PI signals, allowing the identification of viable, apoptotic, and necrotic cells.
RAW 264.7 cells from the initial culture were plated in a 6-well plate and incubated for 24 hours. They then got treated with LPS at a dose of 1 µg/mL and incubated for 24 hours and then harvested. RNA-isolation were conducted by RNeasy plus mini kit (Qiagen). The harvested cell pellets were added 600 µL of RLT plus buffer and the mixture was vortexed for 30 s thoroughly to ensure a complete cell lysis. The mixture was then transferred to the spin cartridge with a collection tube and centrifuged at 12,000×g for 30 s at 4°C. After centrifugation, the flow-through was saved and the 600 µL of 70% ethanol was added. The mixture was gently pipetted and transferred to new spin cartridge. Then the mixture samples were centrifuged at 12,000×g for 15 s and flow-through was discarded. After, 700 µL of RW1 buffer was added to spin cartridge and centrifuged at 12,000×g for 15 s. Then, the flow-through was discarded. 500 µL of RPE wash buffer was added to spin column and centrifuged at 12,000×g for 15 s and the flow-through was discarded and the spin cartridge reinserted into the same collection tube. This process was repeated once and additionally centrifuged at 12,000×g for 2 mn to dry the membrane with bound RNA. After, the flow-through was discarded and the spin cartridge inserted into a recovery tube of 1.5 mL. 30 µL of RNase free water was added to the center of the spin cartridge and centrifuged at 12,000×g for 1mn to elute RNA from the membrane into the recovery tube. RNA quantity was determined using spectrophotometer. RNA measurements obtained were then used to calculate the volume of RNA, H2O, 5X PrimeScript RT Master Mix to be mixed for cDNA synthesis. cDNA synthesis was conducted using PrimeScript RT Master Mix (RR036A, Takara Bio).
To quantitate gene expression levels of Inflammation marker genes and signaling cascade located genes under LPS stimulus and LPS-microbiome co-cultured stimulus, a quantitative real-time polymerase chain reaction (qPCR) was conducted using the BioRad CFX-96 apparatus (BioRad). Sequence-specific primers (Table 1) were designed using Primer-BLAST PRIMER3 software (http://bioinfo.ut.ee/primer3-0.4.0/). Each reaction was carried out in a 20 μL mixture containing 10 μL of TB green Premix Ex taq Ⅱ, 1 μL of forward primer (10 pmoL), 1 μL of reverse primer (10 pmoL), 0.4 μL of ROX reference Dye, 6.6 μL of distilled water, and 1 μL (200 ng/μL) of cDNA. PCR conditions were as follows: a predenaturation step of 94°C for 5 min; 39 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s; and a final step of 72°C for 10 min. All measurements were performed in triplicate for all specimens, and the 2−ΔΔCt method was used for comparing the data. The relative expression of each target gene was calculated by normalizing the expression level against that of glyceraldehyde-3-phosphate dehydrogenase.
RESULTS AND DISCUSSION
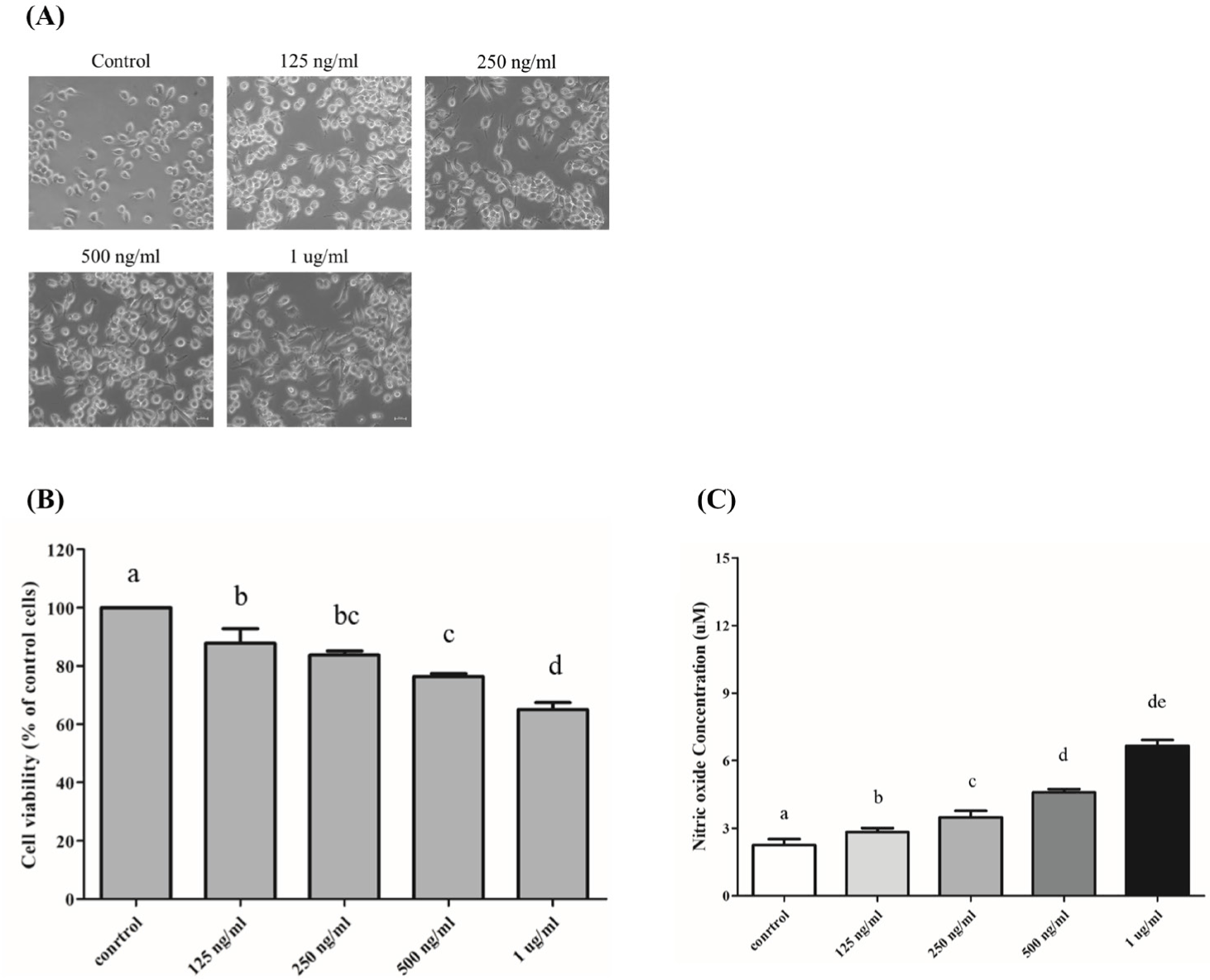
In order to established LPS treatment conditions, RAW 264.7 macrophage were exposure with various concentration (125 ng/mL, 250 ng/mL, 500 ng/mL, and 1 μg/mL) of LPS for 24 hours (Fig. 1A). Significant cytotoxicity was observed in all concentrations (Fig. 1B). The cell viability assays revealed a statistically significant decrease in cellular viability, with a notable drop observed specifically at the concentration of 500 ng/mL of PG-LPS. To determine the certain treatment concentration of LPS on RAW 264.7 macrophage, we conducted NO assay, and mRNA expression analysis with inflammation marker genes. As a result of NO assay, NO production with RAW 264.7 on LPS treatment were increased by showing dose depend expression (Fig. 1C). NO production by RAW 264.7 cells incubated LPS at concentration of control, 125, 250, 500 ng/mL, and 1 μg/mL for 24 hours were 2.25 μM ± 0.23, 2.83 μM ± 0.30, 3.48 μM ± 0.50, 4.59 μM ± 0.24, 6.64 μM ± 0.47, respectively. NO is an essential signaling molecule that plays a critical role in various physiological functions, and it is involved in a wide range of physiological processes, including vasodilatation, increased blood flow, inhibition of platelet aggregation, regulation of inflammatory responses, and cell death [24]. NO, involved in diverse physiological processes, can exert cytotoxic effects when excessively produced. NO has been implicated in cellular and organ dysfunction, as well as oxidative damage. Moreover, NO can promote rapid viral evolution by creating an oxidative stress environment [25]. NO can induce cell cycle arrest or cytotoxicity not only in invading microorganisms but also in the cells that produce it and surrounding cells [26]. Its expression is also reported to be upregulated in response to LPS stimulation [27].
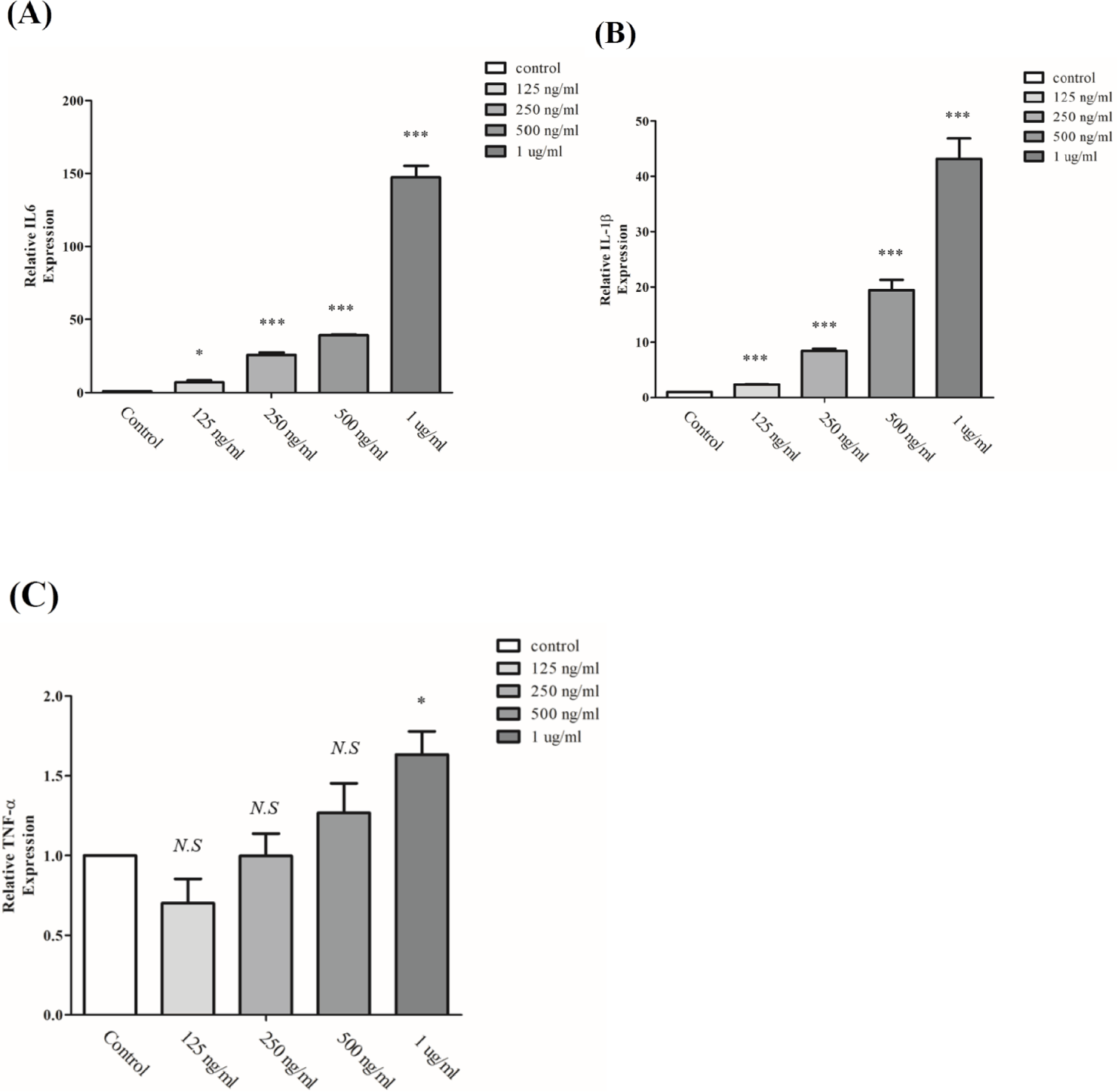
Interferon family genes were normally expressed on immune response [28]. Once infection by pathogen started, LPS induced immune response through Interferon signaling pathway [29]. Those reaction were occurred in a short time. Normally it is determined from 6 hours to 48 hours [30]. Transcripts expression of inflammation markers, IL6, IL1B, and TNF were quantitated by qRT-PCR. The results showed that expression of IL6, IL1B and TNF were significantly increased by LPS dose, even though expression of TNF-α at point of 125 ng/mL were decreased (Fig. 2). IL6 were gradually increased by LPS treatment. At point of 1 μg/mL LPS were approximately 150 times increased compared with control (Fig. 2A). The expression pattern of IL1B revealed a progressively increasing curve, and the expression score at 1 μg/mL LPS was approximately 40 times higher than that of the control group (Fig. 2B). In case of TNF, the growth curve decreased at 125 ng/mL, but LPS treatment from 250 ng/mL to 1 μg/mL affected to RAW 264.7 macrophage proliferation (Fig. 2C). Collectively, the results presented indicate that 1 μg/mL of LPS treatment were highest expression in immune response. Inflammatory responses are typically assessed by evaluating the expression of IL1B, IL6, and TNF genes [28]. The mechanism of inflammation involves a complex cascade of events initiated by cellular damage triggered by external stimuli. This damage leads to the increased expression of pro-inflammatory cytokines, including IL1B, IL6, and TNF. These cytokines, in turn, promote the infiltration of inflammatory cells, which further amplify the inflammatory response by generating reactive oxygen species (ROS) and NO. These reactive molecules activate transcription factors such as NF-κB and COX-2, leading to the upregulation of cell proliferation, apoptosis resistance, angiogenesis, and immunosuppression. These processes collectively contribute to the development of inflammation-induced diseases, including carcinogenesis [31]. In this study, the inflammatory response was assessed by evaluating the expression of pro-inflammatory cytokines, including IL1B, IL6, and TNF.
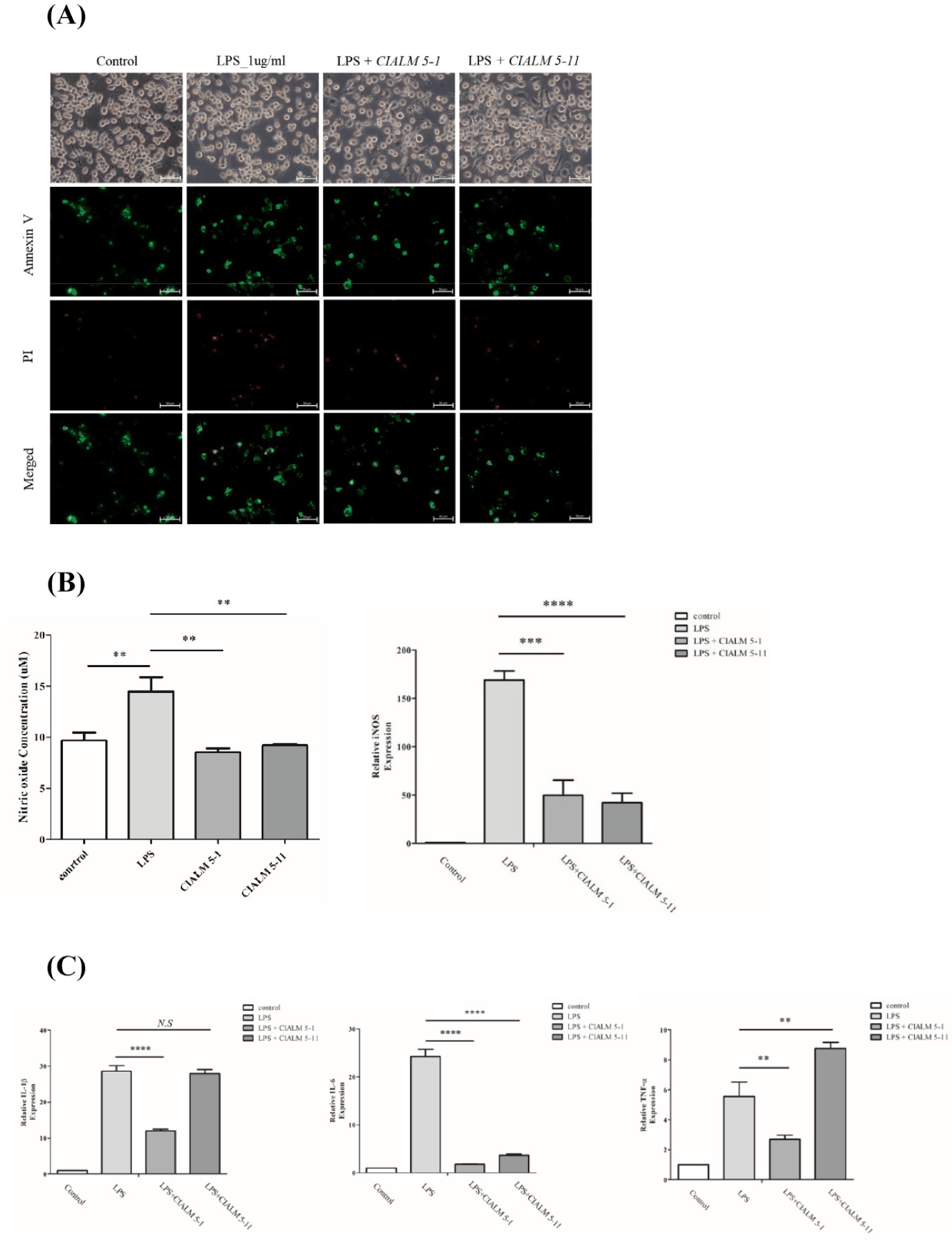
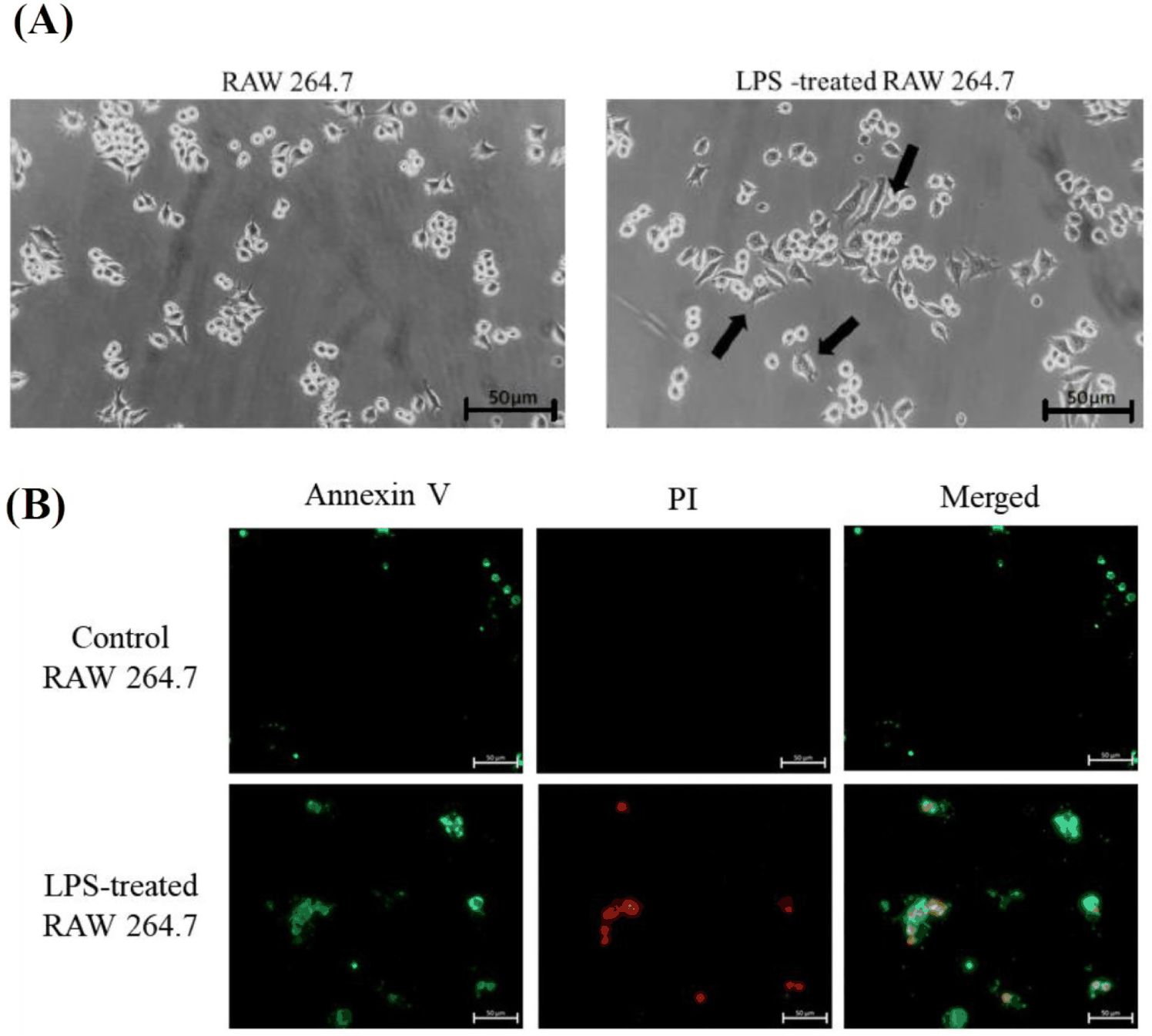
To reconfirm that the established LPS system can induce inflammation in RAW 264.7 macrophages, immune responses were compared between control RAW 264.7 macrophages and those treated with 1 µg/mL LPS (Fig. 3). In a morphological analysis, cytoplasmic vesicles in RAW 264.7 macrophage were observed under LPS treatment (Fig. 3A). Also, annexin V/ Pi staining were conducted and result showed necrosis were slightly increased in LPS treated group than control group (Fig. 3B). Annexin V/PI staining is a widely employed technique for apoptotic cell analysis, comparable to the TUNEL assay. Annexin V is a protein that specifically binds to phosphatidylserine (PS), a phospholipid that is normally localized to the inner leaflet of the cell membrane. However, during apoptosis, PS becomes exposed on the outer leaflet of the cell membrane, allowing Annexin V to bind and label apoptotic cells [32]. Therefore, Annexin V is widely employed for the identification of apoptotic cells. Propidium iodide (PI) stains nuclear DNA in dead cells, as it can enter cells with compromised membranes due to cell death. Because of the mechanism, PI is commonly used in conjunction with Annexin V to identify cell death, and the extent of Annexin/ PI staining allows for the discrimination of early apoptosis, late apoptosis, and necrosis. Collectively, the results of this experiment demonstrate that LPS treatment induces necrosis.
In this study, we collected oral swab samples from two groups of dogs: five healthy dogs and five dogs diagnosed with periodontitis. To identify potential probiotic candidates, we isolated and characterized microorganisms from both groups. Through selective culturing on MRS medium and subsequent 16S rRNA gene sequencing, we identified specific bacterial strains that were significantly more abundant in the oral microbiomes of healthy dogs. These strains were selected as potential probiotic candidates for their potential to mitigate periodontitis.
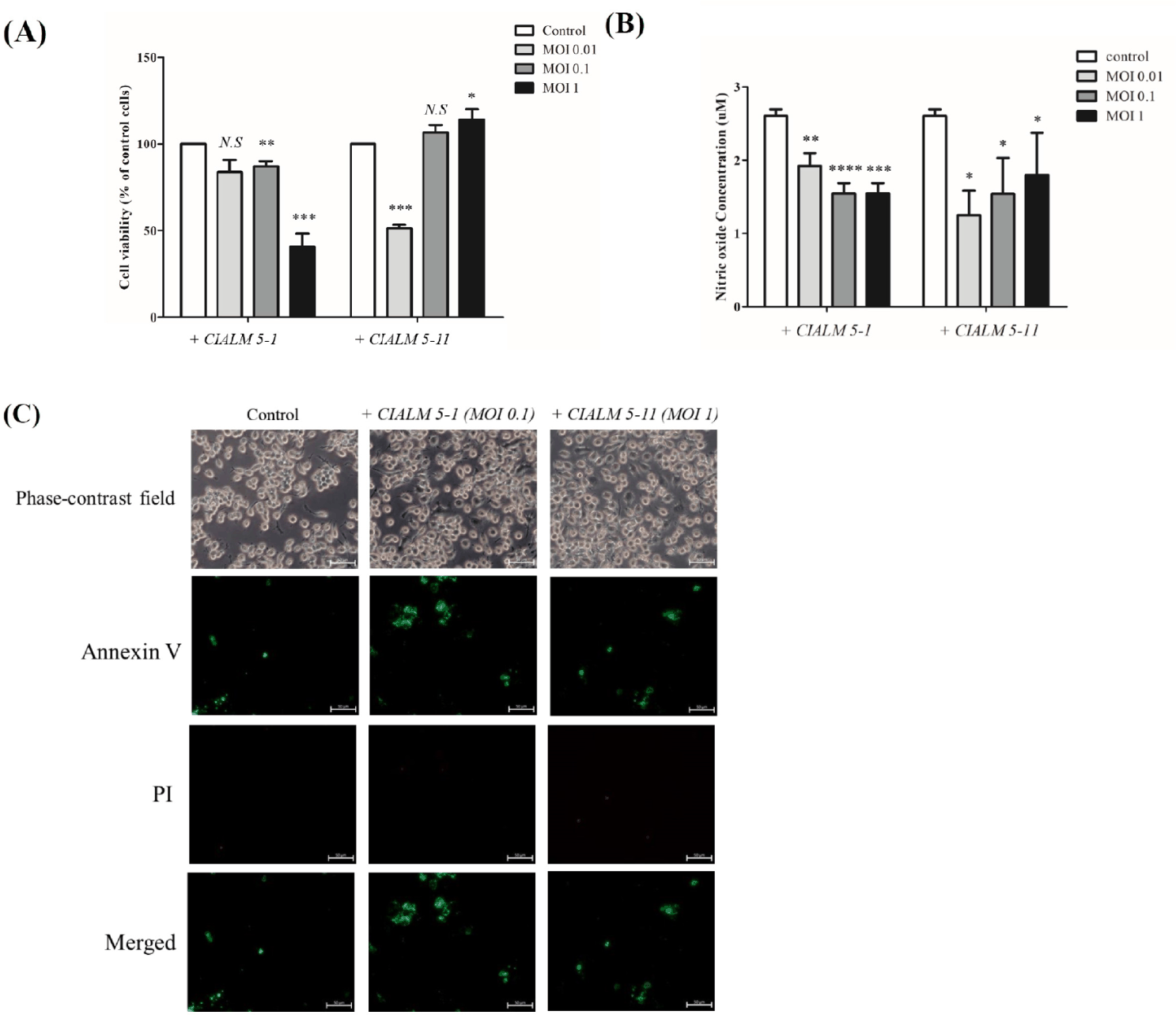
To identify the LPS response reduction effect of C. familiaris canine derived microbiome, cytotoxicity test of the microbiome was conducted with RAW 264.7 macrophage. Cell viability tests were performed on RAW 264.7 macrophages treated with the strains (Fig. 4). Cytotoxicity test were conducted at MOI 1, MOI 0.1, and MOI 0.01 (Fig. 4A). For CIALM 5-1, cell death was significant at MOI 1, but cytotoxicity was not high at MOI 0.1 and 0.01. Interestingly, for CIALM 5-11, cell death was significantly increased at MOI 0.01, but cytotoxicity was not high at MOI 0.1 or MOI 1. In conclusion, each strain showed optimal results for strain efficacy testing at MOI 0.1 for strain CIALM 5-1 and MOI 1 for strain CIALM 5-11. For CIALM 5-11, the reduced cell viability at lower MOI can be explained by the microbial growth cycle. During the exponential growth phase, rapid microbial proliferation leads to the production of substantial amounts of toxic substances [33]. At lower initial inoculum sizes, microorganisms have sufficient time to reach this phase and accumulate enough toxins to compromise cell viability. In addition, Cytotoxicity test of obtained microbiome were carried out with annexin V/ Pi staining. Microbiome labelled as CIALM 5-1, CIALM 5-11 has no significant differences in cytotoxicity compared with control group (Fig. 4B). Nitric Oxide generation (Fig. 4C). Thus, our results demonstrate that the MOIs used for the CIALM strains in the cell viability assays were appropriate concentrations that did not induce cell damage.
To identify the isolated microbiome, 16S rRNA sequencing was performed. The results revealed that CIALM 5-1 identified as Bacillus subtilis showing 99.53% similarity, while CIALM 5-11 showed 99.72% similarity to Bacillus velezensis (Table 2). B. subtilis, a Gram-positive, rod-shaped bacterium, has used as a versatile tool in biological research, alongside the Gram-negative bacterium Escherichia coli. The B. subtilis is found in diverse environments, including soil, air, and water, and is generally considered non-pathogenic [34]. Sharing similar characteristics, B. velezensis is another Gram-positive bacterium. Once considered a subspecies of B. subtilis [35,36], scientific advancements have led to its reclassification as a distinct species in recent years. Similar to B. subtilis, B. velezensis has emerged as a captivating microorganism in recent years, garnering significant interest for its potential applications. This versatile bacterium boasts a remarkable range of functionalities and stability, making it a valuable candidate in areas like biological pesticide [35], fertilizers, and even feed additives [37].
To evaluate the inflammation reduction ability of the obtained strains against LPS, LPS-treated RAW 264.7 macrophages were co-cultured with the strains (Fig. 5). The efficacy of the obtained strains was evaluated using Annexin V/PI staining, NO production assay, and gene expression analysis of inflammatory markers. Annexin V/PI staining revealed that LPS treatment upregulated the expression of both Annexin V and PI. Treatment with candidate microbiome strains CIALM 5-1 and CIALM 5-11 in inflammation induced RAW 264.7 macrophage resulted in a modest reduction in the expression of both Annexin V and PI compared to the LPS-treated group, although the reduction was not significantly reached to control group (Fig. 5A). Similarly, NO production analysis was conducted, and it was found that LPS treatment increased NO production by approximately 50% (Fig. 5B left panel). Subsequently, the effects of candidate strains CIALM 5-1 and CIALM 5-11 on NO production by LPS were investigated. NO production in inflammation induced RAW 264.7 macrophage were significantly suppressed by both of CIALM 5-1 and CIALM 5-11 treatment. Furthermore, analysis of iNOS expression patterns revealed that treatment with both CIALM 5-1 and CIALM 5-11 suppressed LPS-induced iNOS expression (Fig. 5B right panel). Finally, the expression patterns of inflammatory marker genes were evaluated (Fig. 5C). Consistent with the previous experiments, LPS treatment resulted in a dramatic increase in IL-1β expression. treatment of the candidate microbiome strains effectively modulated IL1B expression. Notably, CIALM 5-1 treatment resulted in a significant reduction in IL1B expression, exhibiting a two-fold down-regulation compared to the LPS-treated group. Conversely, CIALM 5-11 treatment did not induce a statistically significant decrease in IL1B expression. Similarly, LPS treatment induced an approximately 25-fold increase in IL6 expression compared with control group. Notably, treatment with both CIALM 5-1 and CIALM 5-11 significantly suppressed IL6 expression, resulting in an approximately 8-fold reduction compared to LPS-treated cells. TNF were significantly two-fold decreased in CIALM 5-1, while CIALM 5-11 showed a slight increase in TNF expression. it is assumed that increase of TNF expression by CIALM 5-11 were enhanced for cell survival and proliferation by activation of JNK/MAPK or PI3K/AKT signaling pathways [38–40]. These findings are consistent with the previously observed cell viability analysis under CIALM 5-11 treatment (Fig. 4B). Research on periodontitis, and oral inflammation has encompassed investigations into oral microbial communities, the activation of the immune system associated with the inflammatory response in periodontal tissues, and the identification of biomarkers for diagnosing and predicting the progression of periodontitis. In recent years, numerous researches were focused on developing novel therapies to mitigate periodontitis and oral inflammation. Therefore, studies have also been conducted on the use of the microbiome to mitigate periodontitis [41,42]. Research on B. subtilis has investigated on the production of antibiotics, the production of enzymes for industrial applications, and the development of biopesticides, plant growth promoters, and feed additives in agriculture and animal husbandry [43]. Investigating the effectiveness of B. subtilis strains isolated from soybean mash and soil-derived Bacillus licheniformis in alleviating periodontal disease were conducted to reduce periodontitis [44]. In addition, researches on efficacy of E-300, isolated and manufactured from the culture supernatant of Japanese soil-derived B. subtilis [45], and B. subtilis-derived VITALREXTM tablets in reducing periodontitis were investigated [46]. Similarly, research on B. velezensis has focused on plant growth promotion, pathogen suppression through antibiotic production, and soil improvement [37]. However, research on the efficacy of B. subtilis and B. velezensis in reducing periodontitis in C. familiaris (dog) remains an area requiring further investigation. This study aimed to identify novel microorganisms for periodontitis reduction by isolating and characterizing microorganisms from the oral cavity of dogs with periodontitis and healthy dogs. The efficacy of B. subtilis and B. velezensis in reducing periodontitis was evaluated through in vitro experiments. In conclusion, both strains were observed to suppress inflammation signaling in RAW 264.7 macrophages, and leading to a reduction in the production of NO, a known inflammatory mediator. The findings of this study provide preliminary data that could inform the development of dietary supplements for reducing periodontitis in dogs, but Further research is warranted to elucidate the molecular mechanisms underlying the periodontitis-reducing effects of these strains and to optimize their cultivation conditions for large-scale production.