
Background
Nutritional, environmental and immune challenges associated with weaning may lead to considerable economic losses to pork producers. This period is generally characterized by decreased voluntary feed intake, altered gut integrity and increased concentrations of inflammatory cytokines in blood [1–3]. These nutritional and physiological abnormalities often result in diarrhea and depression of growth in newly weaned piglets. Restrictions of antibiotic usage in swine have compelled the industry to find alternatives that offer both performance enhancement and protection from disease [4, 5]. In this regard, Liu et al. [6] reported that dietary fish oil reduced the release of pro-inflammatory cytokines in weaned pigs challenged with Escherichia coli lipopolysaccharide. A more recent study indicated that prenatal exposure to long-chain n-3 PUFA increased postnatal glucose absorption in piglets [7]. Although exact mechanisms by which dietary n-3 PUFA modulate immune and metabolic functions in pigs are yet to be fully elucidated, the above study would indicate that dietary n-3 PUFA may help the piglets adapt quickly to the rapidly changing diet at weaning [7].
Currently, there is very little information regarding the use of n-3 PUFA in the diets of pigs raised under minimal disease and stress conditions. To test the hypothesis that nutritional management strategies that attenuate intestinal inflammation may partition nutrients to skeletal muscle for optimal growth, this study was designed to examine the effects of dietary n-3 PUFA on growth and immune response of weanling pigs raised without an added bacterial or environmental challenge.
Results and discussion
Weaning imposes tremendous stress on piglets and is accompanied by marked changes in gastrointestinal physiology, microbiology and immunology [8]. The biochemical and histological changes that occur in the small intestine cause excessive secretion of pro-inflammatory cytokines and induce severe intestinal inflammation. Omega-3 PUFA are known to possess anti-inflammatory properties in humans [9, 10], swine [6, 11] and chickens [12]. To test the hypothesis that nutritional management strategies that attenuate intestinal inflammation may repartition nutrients to tissue accretion, we examined the effects of dietary n-3 PUFA on growth and immune response of weanling pigs (Figure 1) raised without an added bacterial or environmental challenge.
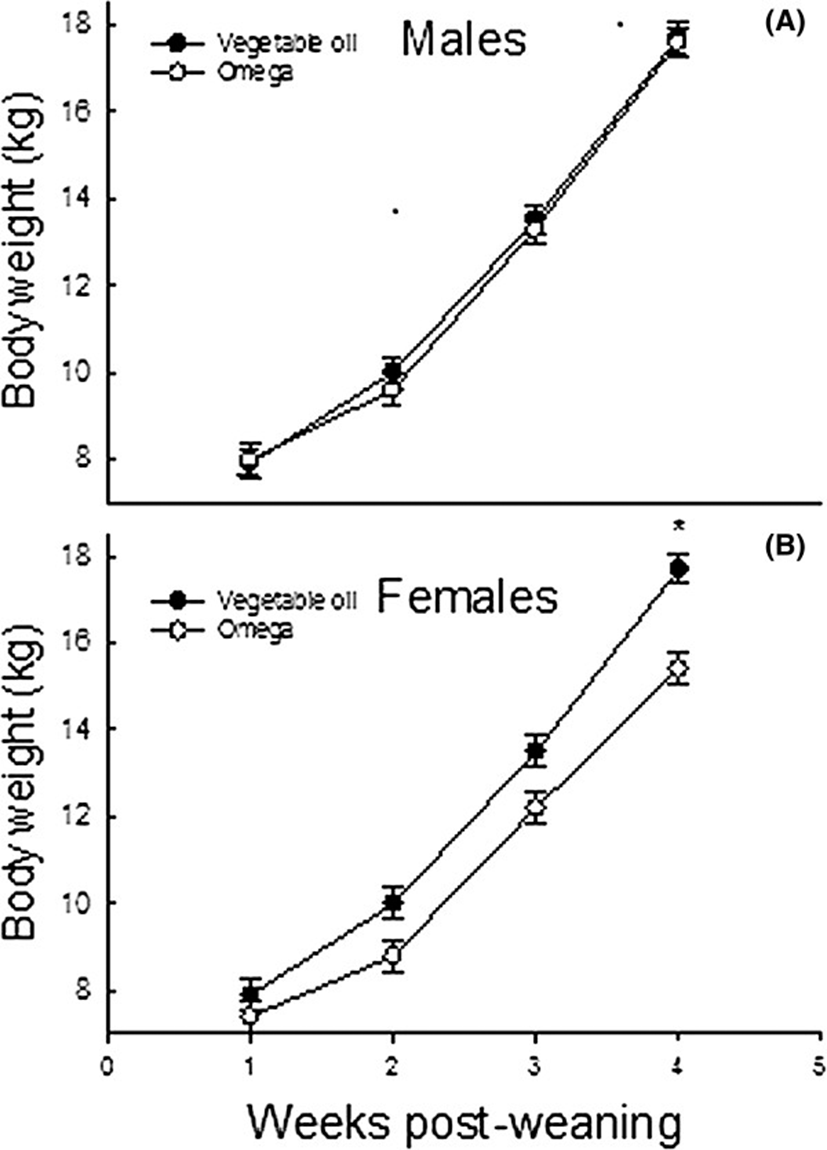
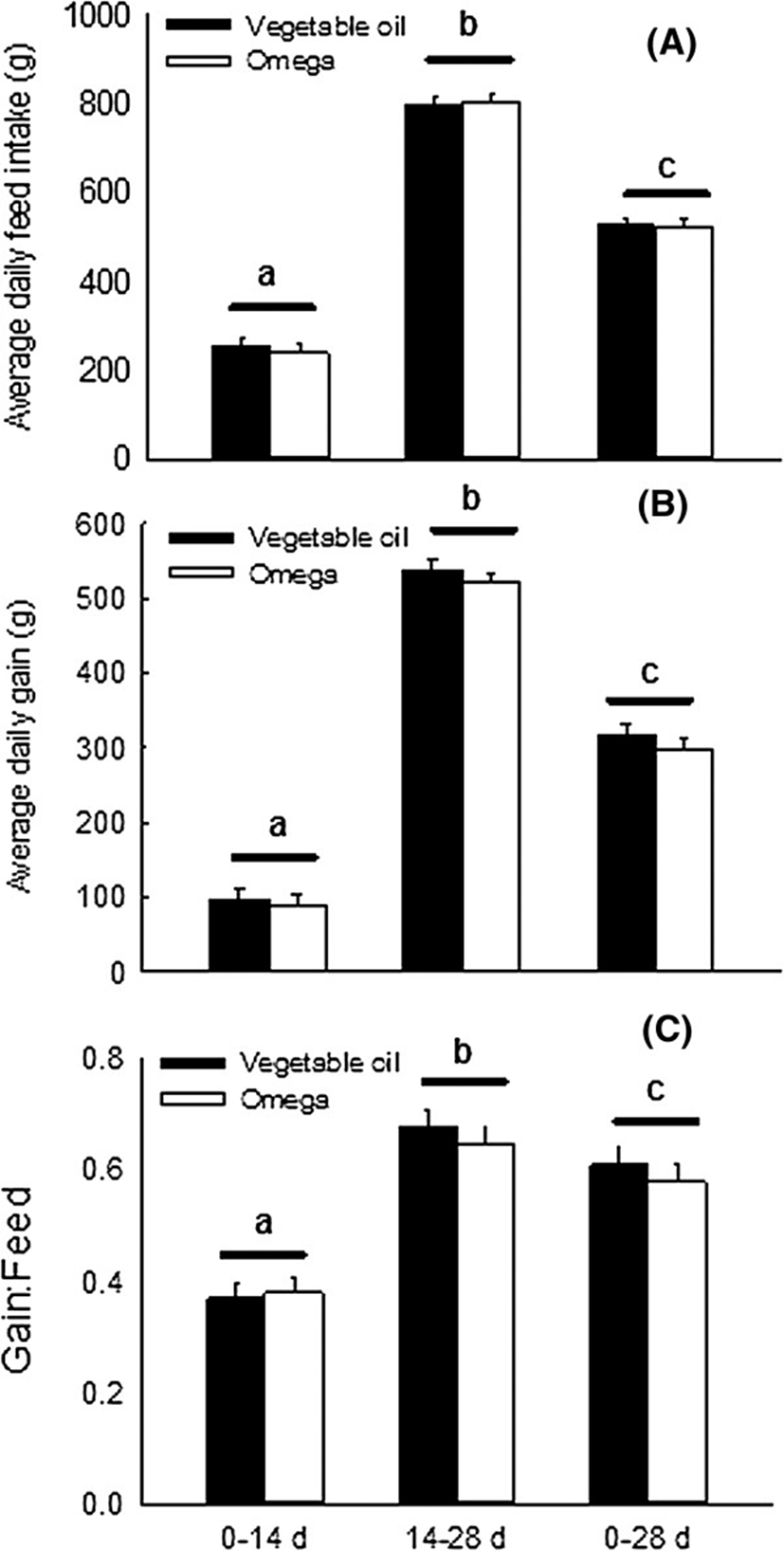
Inclusion of 3% n-3 PUFA in the weanling piglet’s diet did not result in significantly improvement of average daily gain (ADG), average daily feed intake (ADFI) or growth to feed ratio (G: F) in weanling pigs. These findings are consistent with an earlier study [13] which detected no effects of dietary flax seed meal (rich in alpha linolenic acid) on basal body weight gain, feed intake or feed efficiency in weanling pigs. Additional studies using control diets with less 18: 3n-3 are needed to examine true effects of long-chain n-3 PUFA on growth and feed intake responses. Female piglets consuming the n-3 PUFA-supplemented diet were lighter at week 4 post-weaning than those consuming the vegetable oil-enriched diet. Whether or not this phenomenon was due to alteration in body composition as a result of feeding n-3 PUFA to nursery pigs was not documented. In rodents [14–16] and humans [17, 18], diets rich in n-3 PUFA lower fat stores and increase lean tissue mass. It is, therefore, possible that the smaller body weight of female piglets consuming n-3 PUFA detected at week 4 post-weaning was due to a decrease in fat accretion at the expense of lean tissue. The improvement of body weight gain detected in experimental animals between days 14 and 28 post-weaning likely resulted from an increase in feed intake and a decrease in basal inflammatory challenges during the second phase of growth (Figure 2).
Concentrations of IGF-I in plasma decreased immediately following weaning and increased again by day 28 post-weaning (Figure 3). These findings indicate that weaning may cause a significant metabolic stress in weanling pigs and that this stress decreases with increasing weeks after weaning. There is little information on the effect of dietary n-3 PUFA on peripheral concentrations of growth factors in the pig. In the present study, inclusion of 3% n-3 PUFA into the piglet’s diet had no detectable effects on plasma IGF-I concentration during the first four weeks after weaning. These observations are consistent with previous studies [6, 13] which showed no beneficial effects of dietary fish oil on basal IGF-I concentration in weaned pigs. Thissen and Verniers [19] reported that IL-6 and TNF-α decreased both growth hormone (GH) and IGF-I mRNA in rat hepatocyte primary cultures. We did not examine GH or IGF-I transcript modulation by inflammatory cytokines, and therefore, whether or not the lack of n-3 PUFA effects on plasma IGF-I concentration detected in the present study was indicative of cytokine-mediated uncoupling of GH and IGF-I gene expression in weanling pigs warrants further investigation.
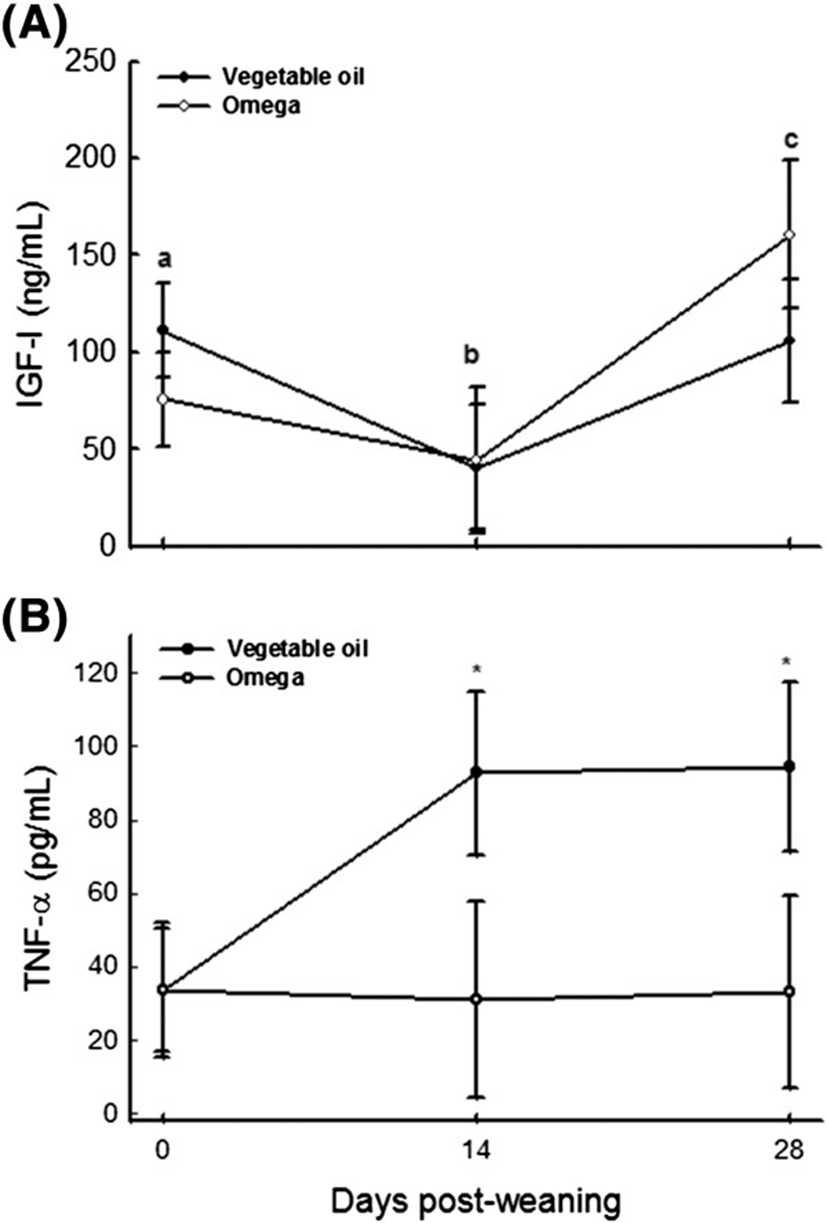
Tumor necrosis factor-α, a cytokine produced primarily by monocytes and macrophages, is thought to be one of the principal mediators of inflammation [20]. In the present study, plasma TNF-α concentrations were lower in weanling piglets supplemented with n-3 PUFA than those fed the vegetable oil supplement (Figure 3). These findings are consistent with previous in vitro [21–23] and in vivo [11, 24, 25] studies and suggest that n-3 PUFA inclusion in the diet could mitigate the immune stress in weanling pigs. Whereas exact mechanisms of n-3 PUFA suppression of TNF-α are yet to be fully elucidated, we speculate that suppression of TNF- α production by n-3 PUFA may be attributed, in part, to their inhibitory effects on NF-κB activation and or translocation to the nucleus [9, 22, 23]. Nuclear factor-κB are normally confined in the cytoplasm through their association with IκΒ. When cells are activated by inflammatory stimuli, the IκΒ are rapidly phosphorylated and degraded to free the NF-κΒ. The free NF-κΒ then migrate to the nucleus where they bind to cognate DNA binding sites and activate inflammatory gene transcription [9]. Any factor that prevents IκΒ phosphorylation and, thus, NF-κΒ activation, will decrease pro-inflammatory gene expression in the nucleus. Additionally, long-chain PUFA serve as ligands for peroxisome proliferator-activated receptors (PPAR), which are known to inhibit nuclear translocation of NF-κΒ [9]. Thus, activation of PPAR may be another intracellular mechanism by which marine n-3 PUFA regulate NF-κΒ activation and TNF-α production in animal models [9].
Hematological traits of swine are influenced by a variety of environmental and physiological factors including diet, age, gender and housing [26, 27]. In the present study, most of the blood characteristics examined did not differ among pigs fed the two diets (Table 1). Blood samples for complete blood cell counts were collected at 4 weeks after weaning, and it is possible that by this sampling time, the weanling piglets had already recovered from most physiological and dietary challenges normally associated with weaning in pigs. Alternatively, the piglets used in this study were raised in a clean environment and, thus, may not have acquired the “normal” gastrointestinal microflora that would cause clinical diseases. This hypothesis was further supported by our inability to detect salmonella and enterotoxigenic E. coli in fecal samples collected at week 4 post-weaning (data not shown).
aMeans represent 4 pigs per dietary treatment.
bDiets were: Control (3% vegetable oil) and omega (3% Gromega Ultra 345, provided by JBS United, Inc., Sheridan, IN).
cP-values for control compared to Omega diet.
dWhite blood cells.
eRed blood cells.
Conclusions
In the pig, the period following weaning is generally characterized by sub-optimal growth, deteriorated feed efficiency, and a high incidence of diarrhea. Results of this study provided no evidence for n-3 PUFA modulation of growth of male weanling pigs raised in the absence of significant immunological and environmental challenges. The observation that female piglets consuming the n-3 PUFA-supplemented diet were lighter at week 4 post-weaning than those consuming the vegetable oil-enriched diet (Figure 2) may be indicative of a decrease in fat accretion at the expense of lean tissue. Additionally, dietary n-3 PUFA may improve the immune status of weanling pigs, as reflected by considerably lower plasma TNF-α in pigs consuming n-3 PUFA than those fed vegetable oil. The gradual increase in body weight, feed intake and feed efficiency following weaning likely reflects a progressive adaptation to post-weaning diets and a gradual improvement of the gastrointestinal microbiota.
Methods
The animal protocol for this research was approved by the institutional Animal Research Committee of the University of Florida. To avoid potential differences due to farrowing season, the study was conducted using 40 piglets born within one week at the Swine Research Unit of the University of Florida (Gainesville, FL) during the month of March 2013. Forty crossbred pigs (averaging 27 ± 2 days of age and 8.1 ± 0.7 kg of body weight) were balanced for initial body weight and gender across two treatment groups in a complete randomized block design. Experimental animals were fed either a control (3% vegetable oil, n = 20) or n-3 PUFA (3% marine n-3 PUFA; Gromega 345, JBS United, Inc, Sheridan IN, n = 20)-supplemented diet for four weeks after weaning. The vegetable oil was purchased from Sysco Corporations (Houston, TX) and contained approximately 22% total fat. Omega-3 fatty polyunsaturated fatty acids used in this study were provided by JBS United (Sheridan, Indiana) and contained a minimum of 39% crude fat. Complete ingredient compositions and FA profiles of experimental diets are summarized in Tables 2 and 3, respectively. Pigs were housed in pens (groups of 5 animals per pen; pen size = 2.4 m × 1.8 m) and kept on the same diet for the entire experimental period. Body weight and feed consumption were recorded weekly throughout the 4-week experiment. These observations were used to calculate ADG, ADFI, and G: F.
aDiets were: Control (3% vegetable oil) and omega (3% Gromega Ultra 345, provided by JBS United, Inc., Sheridan, IN).
aFatty acid analysis was performed by the University of Missouri Analytical Laboratory.
bDiets were: Control (3% vegetable oil) and omega (3% Gromega Ultra 345, provided by JBS United, Inc., Sheridan, IN).
On days 0, 14 and 28 of the experiment, jugular venous blood samples (8 ml from each experimental pig) were collected into evacuated heparinized tubes (BD Franklin Lakes, NJ) and centrifuged (3,000 × g for 15 min) to separate plasma. The plasma samples were stored at −80°C until analysis. Concentrations of IGF-I and TNF-α in plasma were analyzed using commercially available ELISA kits (R&D Systems, Inc., Minneapolis, MN). Hormone and cytokine analyses were performed in single assays and intra-assay CV were 4.0 and 4.7% for IGF-I and TNF-α, respectively. The least detectable concentrations were 0.06 ng/mL and 5.50 pg/ml. On day 27 of the experiment, additional blood samples were collected for complete blood cell counts, and hematological traits were determined as described by Quiroz-Rocha et al. [28].
Two fecal consistency scores were assigned to each pen on weeks 1, 2, 3, and 4 post-weaning. The scale used to assess fecal consistency was based on a numerical scale of 1 to 3, where 1 represented a normal (hard) feces, 2 represented a soft (moist) feces, and 3 represented diarrhea (watery liquid). The weekly score for each pen was calculated by averaging the two fecal consistency scores (Table 4).
| Week post-weaning | Dietsc | Omega | SEM | Pd |
|---|---|---|---|---|
| Control | ||||
| 1 | 2.6 | 2.5 | 0.2 | 0.67 |
| 2 | 1.9 | 1.9 | 0.2 | 1.00 |
| 3 | 2.0 | 1.9 | 0.2 | 0.68 |
| 4 | 1.3 | 1.1 | 0.2 | 0.68 |
aThe scale used for assessing fecal consistency was based on a numerical scale of 1 to 3, where 1 represented a normal (hard) feces, 2 represented a soft moist feces, and 3 represented diarrhea (watery liquid).
bMeans represent average fecal scores for 4 pens per dietary treatment.
cDiets were: Control (3% vegetable oil) and omega (3% Gromega Ultra 345, provided by JBS United, Inc., Sheridan, IN).
dP-values for control compared to Omega diet.
Effects of diets on growth, IGF-I, TNF-α and fecal characteristics were analyzed using the MIXED procedure of Statistical Analysis System (version 9.3) with repeated measures [29]. For individual measurements (body weights), fixed effects included diet, gender, diet × gender interaction, week after weaning, diet × week interaction, gender × week interaction and diet × gender × week interaction. The pig, nested within gender and diet, was considered a random variable, and therefore the pig variance was used to test the effects of diet, gender, and diet x gender interaction. Initial weights were used as covariates in these analyses. A similar model was used to test the effect of diet on plasma IGF-I and TNF-α concentrations, except that week after weaning was replaced by day of blood sample collection. For collective measurements (feed intake, average daily gain, feed efficiency, and fecal consistency score), the statistical model included the effect of diets, pen (diet), week relative to weaning, diet × week interaction. In these models, pen was used as experimental unit to test the main effect of diet. Single blood samples were collected for complete blood cell counts, and, therefore, the statistical models for hematological traits contained only the main effect of diet. For all responses, significant differences between means were declared at P < 0.05.