
INTRODUCTION
Deer antler velvet is a non-osteolytic agent that is widely used in traditional medicine because it contains a variety of small ingredients (e.g., monosaccharides, uronic acid, free amino acids, sialic acid, gangliosides, and small peptides) [1] that promote recovery from fatigue [2], improve liver function and mitigate skeletal weakness symptoms [3,4], strengthen immunity [5], and reduce inflammation. As the elderly population is increasing, demand for health supplements and import of antlers velvets have increased rapidly in recent years. New Zealand, Russia, Canada, and China are countries importing deer antlers, with New Zealand importing greatest quantity and Russia importing the most expensive ones [6]. In Asian countries, deer antler velvets are dried and sold as slices or usually boiled with various types of herbs, botanical materials, and other ingredients in water to be consumed as medicinal soup. Despite having numerous beneficial ingredients, there are limited ways to consume deer antlers. Therefore, there have been various attempts to apply fermentation along with herb materials in order to increase the physiological efficacy of antler velvet and herbal medicine [7].
Microbial fermentation is used in the food industry for a variety of purposes, including the enhancement of flavor, preservation, and health benefits [8]. It is also used to generate useful substances through the enzymatic decomposition of organic matter. During fermentation, high-molecular weight substances are converted into low-molecular weight substances, such as organic acids and amino acids, and glycosides are broken down so that nutrients can be more easily absorbed and utilized by the body [9]. Previous studies have reported that Bacillus subtilis-fermented antler extract promotes hematopoiesis [10], inhibits osteoclast differentiation [11], and improves exercise endurance [12].
In this study, we focused on the optimal fermentation of deer antlers using probiotic strains, mainly the lactic acid bacteria (LAB). Fermentation with Lactobacillus sp. has been reported to enhance absorption of bioactive molecules, and aid in immune modulation, cholesterol homeostasis, exercise performance [13–16]. The purpose of the present study was to enhance the concentration of bioactive ingredients (e.g., sialic acid and gamma-aminobutyric acid [GABA]) in fermented antler velvet (FAV) and evaluate its effects on immune response in Caenorhabditis elegans and mouse models.
MATERIALS AND METHODS
The fer-15;ferm-1 mutants were synchronized using an alkaline hypochlorite solution. Lactobacillus rhamnosus GG (LGG) and Escherichia coli OP50 (OP50) were used as controls. Concentrated suspensions of LAB or E. coli OP50 (100 μL) were plated on 35-mm diameter nematode growth medium (NGM) agar plates. Day 0 was defined as the day of L4-to-adult switch. Worms were transferred, using a platinum wire, onto fresh plates daily and maintained at 25°C. The experiment was repeated three times, and survival rates were determined.
Water extracts of deer antler, Cervus canadensis Erxleben, were procured from Korea RND Nonghyup (Seoul, Korea) and stored at 4°C. To ferment with LAB and antler extract, minimal prebiotic medium with 2% antler extract (1 L standard: 20 g antler water extract, 5 g peptone, 2.5 g sodium acetate, 0.5 mL of 1 M magnesium sulfate heptahydrate, 0.5 mL of 1 M manganese sulfate tetrahydrate, 5 mL Tween 80, 1 g diammonium citrate, 1 g dipotassium phosphate, 20 g glucose, and 1 g L-glutamic acid) was prepared and autoclaved at 121°C for 15 min. The working antler medium was aliquoted into 50-mL conical tubes, and 1% of each of the 17 LAB strains was then inoculated (>104 colony-forming unit [CFU]/mL). After incubation at 37°C for 24, 48, 72, 96, and 120 h, the cultures were centrifuged at 4,000×g for 10 min at 25°C, and the resulting supernatants were filtered using 0.22-µm syringe filters (Minisart, Sartorius Stedim Biotech GmbH, Goettingen, Germany).
The acidity of each culture (1 mL) was measured by adding 10 µL of 1% phenolphthalein and then adding 0.1 N NaOH until reaching an endpoint. The amount (mL) of 0.1 N NaOH consumed was then converted and expressed as lactic acid content (%). The culture pH was measured during fermentation using a pH meter (S20 SevenEasy pH meter, Mettler-Toledo, Columbus, OH, USA). The pH and acidity of the cultures were measured before and after each fermentation period (24, 48, 72, 96, and 120 h). Bacterial concentration was assessed using the CFU counting method. Briefly, serial diluents (100 µL) were spread on deMan, Rogosa, and Sharpe (MRS) agar plates, and CFUs (Log10/mL) were counted after 2 d.
Total nitrogen content was analyzed photometrically as described previously [17], with minor modifications. Briefly, samples of each culture were diluted 25-fold, mixed with 5 mL aqueous HCl (0.2%), 20 mL ddH2O, and 10 mL potassium persulfate (3% alkalinity, pH 9.7); autoclaved at 120°C for 30 min; cooled to room temperature; and then combined with 5 mL HCl (0.06%). Finally, the absorbance of each mixture was measured at 220 nm, and nitrogen content was determined using a calibration curve generated using potassium nitrate. To measure total amino acid content, amino acids were extracted as described by Sun et al. [3], with slight modifications. Briefly, 50 μL of each culture was mixed with 2 mL ddH2O, 1 mL acetic acid-sodium acetate buffer solution (pH 6.0), and 1 mL aqueous ninhydrin (2%); incubated at 100°C for 15 min; cooled to room temperature; and then mixed with 10 mL 50% ethanol. Finally, the absorbance of each mixture was measured at 220 nm [3], and amino acid concentration was determined using a calibration curve generated using an arginine standard. The protein content of each culture was quantified using Bradford protein analysis, following the manufacturers’ instructions. The concentration of each mixture was determined by measuring absorbance at 595 nm.
GABA analysis was performed according to the method described by Lü et al. [18], with slight modifications. Briefly, 3 mL of each sample was mixed with 1.5 mL NaHCO3 (0.5 M) and 0.5 mL 1-fluoro-2,4-dinitrobenzene (0.715 mg/mL), incubated at 60°C for 1 h, cooled to room temperature, and then passed through a 0.22-μm syringe filter (PTFE, Advantec, Tokyo, Japan). The GABA content of each sample derivative (10 μL) was then analyzed using a Shimazhu 20A series HPLC system (Shimadzu, Kyoto, Japan) equipped with a Symmetry C18 column (5 μm, 3.9 × 150 mm; Waters, Milford, MA, USA). Isocratic elution was performed with 0.5% ammonium acetate aqueous solution and acetonitrile (85:15, v/v) for 20 min at a flow rate of 1 mL/min. GABA was detected based on absorption at a wavelength of 360 nm.
Each sample (0.1 mL) was mixed with 0.1 mL H2SO4 (0.2 N), hydrolyzed at 85°C for 1 h, and then reacted with 20 µL periodate solution (0.25 M NaIO4 in 9 M H3PO4) at room temperature for 20 min. Next, 0.1 mL arsenite reagent (10% NaAsO2 in 0.1 N H2SO4 with 0.5 M Na2SO4) was added to reduce the excess periodate. Thereafter, 0.25 mL thiobarbituric acid (TBA) solution (0.6% TBA in 0.5 M Na2SO4) was added and color was developed at 85°C for 15 min, followed by cooling at room temperature. Finally, each sample was mixed with 1 mL cyclohexanone and centrifuged at 900×g for 7 min. The absorbance of each supernatant was measured at 549 nm (Synergy HTX multi-mode plate reader, BioTek, Winooski, VT, USA) [19], and sialic acid content was calculated by comparison to a standard curve for N-acetyl neuraminic acid.
The supernatants of the fermented samples were collected after centrifugation at 5,500×g for 5 min and filtered using 0.2-μm cellulose acetate syringe filters. Each sample was then mixed with ice-cold methanol, vigorously vortexed on ice for 1 min, and centrifuged at 10,000×g for 10 min at 4°C. The resulting supernatants were filtered using 0.2-μm polyvinylidene fluoride (PVDF) syringe filters (Whatman, Maidstone, UK) and completely vacuum-dried.
For gas chromatography-mass spectrometry (GC-MS) analysis, each sample was dissolved in 30 µL methoxyamine hydrochloride (Sigma-Aldrich, St. Louis, MO, USA) in pyridine (20 mg/mL), derivatized at 30°C for 90 min. Samples were then trimethylsilylated by adding 50 µL N,O-bis(trimethylsilyl)trifluoroacetamide (Sigma-Aldrich) and incubated at 60°C for 30 min. GC-MS analysis was performed using a TRACE™ 1310 Gas Chromatograph (Thermo Fisher Scientific, Waltham, MA, USA) equipped with an ISQ LT single quadrupole mass spectrometer (Thermo Fisher Scientific), and separation was performed using a DB-5MS column (60 m × 0.25 mm, 0.25-um film thickness, Agilent, Santa Clara, CA, USA). Oven temperature was maintained at 50°C for 2 min, gradually increased at 5°C/min, maintained at 180°C for 8 min, gradually increased at 2.5°C/min to 325°C, and then maintained for 10 min. The sample was injected at 300°C, and helium was used as the carrier gas, with a flow rate of 1.5 mL/min and split ratio of 1:60. An electron ionization system with an ionization voltage of 70 eV was used for GC-MS detection, and the ion source temperature was 270°C. The mass scan range was set at 30–450 (m/z), and the acquisition rate was 5 spectra/s. Detected metabolites were identified using the NIST Mass Spectral Search Program (version 2.0; National Institute of Standards and Technology [NIST], Gaithersburg, MD, USA).
The C. elegans killing assay was performed as previously described [20]. L4/young adult worms were placed on conditioning NGM plates with non-fermented antler velvet (AV) medium (10 mg/mL) or FAV (10 mg/mL), and non-pathogenic E. coli OP50 at 25°C for 24 h. Worms were plated at 30 per plate. Subsequently, for viability analysis, nematodes preconditioned with AV, FAV, and OP50 were transferred onto Salmonella typhi SL 1344, Listeria monocytogenes EDG-e, and E. coli O157:H7 EDL933 culture plates; incubated at 25°C; and examined using a microscope at 24-h intervals until all worms died. Whether worms were alive was assessed by gentle touching with a platinum wire.
Spleen tissues were extracted from C57BL/6 mice (24-wk-old female, 20–22 g). The tissues were disaggregated via passage through a 40 µm cell strainer in RPMI-1640 medium (SH30027, Hyclone, Logan, UT, USA) containing 10% fetal bovine serum (FBS), 1% antibiotics and the cells were purified by centrifugation at 1,000×g for 10 min at 4°C. Red blood cells were removed by incubating purified cells in Red blood cell lysis buffer (A10492-0, GIBCO, Thermo Fisher Scientific) for 3 min. Splenocytes were then washed with RPMI containing 10% FBS and 1% antibiotics and centrifuged at 1,000×g for 10 min at 4°C for 10 min.
The cell viability assay was performed using an EZ-Cytox cell viability assay kit (EZ-3000, DoGen Bio, Seoul, Korea). Splenocytes were seeded into 96-well plates (approximately 5×106/mL) and treated with 0, 0.1, 0.5, and 1 mg/mL of the AV or FAV samples for 48 h. Cells were co-treated with 1.6 mg/mL cyclophosphamide (Cy; C0768-1G, Sigma-Aldrich) and the antler velvet sample. Subsequently, 10 µL of EZ-Cytox reagent was added to each well and incubated for 1 h. Absorbance of the samples was then measured at 450 nm. Cell viability values were normalized to those of the cells from the untreated control groups.
Ten-week-old female C57BL/6 mice (19–20 g) were housed in a room with controlled temperature (22 ± 2°C) and humidity (50 ± 5%) on a 12/12 h light/dark cycle with free access to food and water. After 1 wk of acclimatization, the mice were randomly divided into four groups (n = 5): control group, Cy group (100 mg/kg), AV 2 mg/20 g group, FAV 2 mg/20 g group. The AV or FAV-treated groups were orally administered the corresponding concentration of antler velvet for about 28 d, and the control and the Cy groups received oral administration of the same amount of saline. Mice were administered intraperitoneal injection of Cy (100 mg/kg) in a total volume of 200 µL of saline on days 21 and 24. All of the mice were administered with Cy except the control group. At the end of the experiment, mice were sacrificed, and the spleens were collected.
Total RNA was isolated from the spleen tissues using the total RNA Mini Kit (T2010S, New England BioLabs, Ipswich, MA, USA), as per the manufacturer’s instructions. A cDNA kit (170-8891, Bio-Rad Laboratories, Hercules, CA, USA) was used to synthesize cDNA from 1 µg of total RNA, using the primers. Target gene expression was analyzed using the gene-specific primers listed in Table 1. The amplified products were subjected to electrophoresis on a 1% agarose gel and visualized with ultraviolet illumination. For qPCR, cDNA strands synthesized from the isolated total RNA were used as templates. PCR was performed on a StepOnePlus Real-Time PCR System (Applied Biosystems, Waltham, MA, USA), using the SYBR Green PCR Mastermix (1725121; Bio-Rad Laboratories). The cycle threshold values were normalized against the β-actin gene expression levels.
Data were analyzed using GraphPad Prism software (Prism 9 Academic), and the results are presented as mean ± SD. Statistical analysis was performed using Student’s t-test for comparisons between two groups and one-way analysis of variance for comparisons among multiple groups. p < 0.05 was considered significant.
RESULTD AND DISCUSSION
Previously, we had isolated approximately 200 strains of LAB, including L. rhamnosus and L. plantarum strains, from fermented Kimchi and infant feces. Among the isolates, we screened the strains having basic probiotic properties for further fermentation with antler velvet extract. Of the 200 strains isolated from kimchi and infant feces, 17 Lactobacillus strains (L. sakei, L. brevis, L. rhamnosus, and L. plantarum) were selected based on their superior acid resistance, bile resistance, and intestinal adhesion, as shown in Table 2. All the isolates were identified by 16S rRNA sequencing. The study of the selection of 17 strains as suitable candidates for deer antler extract fermentation is unpublished. To evaluate the activity of strains during fermentation with antler extract, 2% of velvet antler water extracts (water extract 50% w/w) containing minimal prebiotic media was used for fermentation. During the first three days of fermentation, all 17 strains maintained sufficient cell populations, lowered culture pH, and increased culture acidity (Table 3). The MR ID stains from infant feces exhibited more rapid pH lowering and higher acidity rather than LFR ID stains from Kimchi, whereas higher cell population was maintained during fermentation with Kimchi strains except for LFR20-006. There have been a few studies on the strengthening of bioactive ingredients of deer antlers via fermentation [10,12,21]. Kim et al. [21] utilized 11 Bacillus sp., 4 Lactobacillus sp., and 6 mushroom stains (Cordyceps militaris) to evaluate the growth in antler medium. All Lactobacillus sp. (L. plantarum, L. mesenteroides, L. delbrueckii) failed to grow in both antler extract and antler power-based media. Surprisingly, all the strains in the present study successfully exhibited high proliferation with 108–109 CFU/g, indicating that all strains were compatible with antler extract and proliferated and produced strain-specific metabolites.
We attempted to evaluate the longevity-promoting effects of all 17 selected strains using C. elegans to identify optimal strains for fermentation. Strains LFR20-003, LFR20-004, LFR20-007, LFR20-008, MR4, MR5, MR14, MR19, MR43, MR45, and MR50 were as efficacious if not more than the positive control LGG (Fig. 1). In our previous study, Lactobacillus plantarum JDFM216 was selected, and it was reported that LAB strains could mitigate immune function decline and cognitive decline due to aging [22,23]. As an important industrial application of LAB, lactic acid fermentation continues to be a highly desirable method for processing and preserving non-dairy foods owing to its low cost, low energy requirements for both processing and preparing foods for consumption, high yield, and diversified flavors as compared to fermentation with mold or Bacillus sp. [24].
On the second day after fermentation with antler velvet, the amounts of total nitrogen, amino acid, proteins, and bioactive substances were measured. In most fermented extracts, the amounts of total nitrogen, protein, and amino acids decreased (Table 4). In general, Lactobacillus has the ability to hydrolyze proteins present in the medium, and this proteolytic activity produces a variety of peptides, metabolites with biological activity, as well as free amino acids required by bacteria. [25,26]. In this experiment, it was found that even for the same Lactobacillus species, the protein utilization rate was variable. This result indicates the diversity in mechanisms involved in the longevity-promoting properties in C. elegans.
LAB producing and releasing GABA, a neurotransmitter, have been reported, and their mechanisms were reviewed [27–29]. Although there are reports of the effect of deer antler extract [30] and bone extract [31] on memory improvement in scopolamine-induced memory impaired mice, the production of GABA in deer antler extract after fermentation has not yet been investigated. The GABA content of the antler velvet was increased six times when fermented by MR43, MR45, and MR50, which were all isolated from infant feces, and was increased three and two times when fermented by LFR20-007 and LFR20-004, respectively, which were isolated from kimchi (Fig. 2A). In addition, the amount of ganglioside, a major bioactive component of deer antlers, was measured. Sialic acid in antler velvet has been considered a biomarker among bioactive molecules. Various extraction methods utilizing enzymes and fermentation, amongst others, have been developed to improve the recovery of sialic acid [32–34]. All the selected strains increased the sialic acid content; however, fermentation with LFR20-006, LFR20-007, and LFR20-008 increased the sialic acid content by approximately eight-fold (Fig. 2B). Interestingly, after fermentation, the levels of sialic acid, rather than GABA, in most fermented extracts were increased dramatically. Park et al. [10] reported that the sialic acid content was highest, exhibiting a four-fold increase approximately in Bacillus subtilis-FAV extract on the fourth day. The 11 LAB strains in this study (LFR20-001, -003, -004, -005, -006, -007, -008, -009, MR4, MR5, and MR19) exhibited greater ability to produce sialic acid in fermented deer antler.
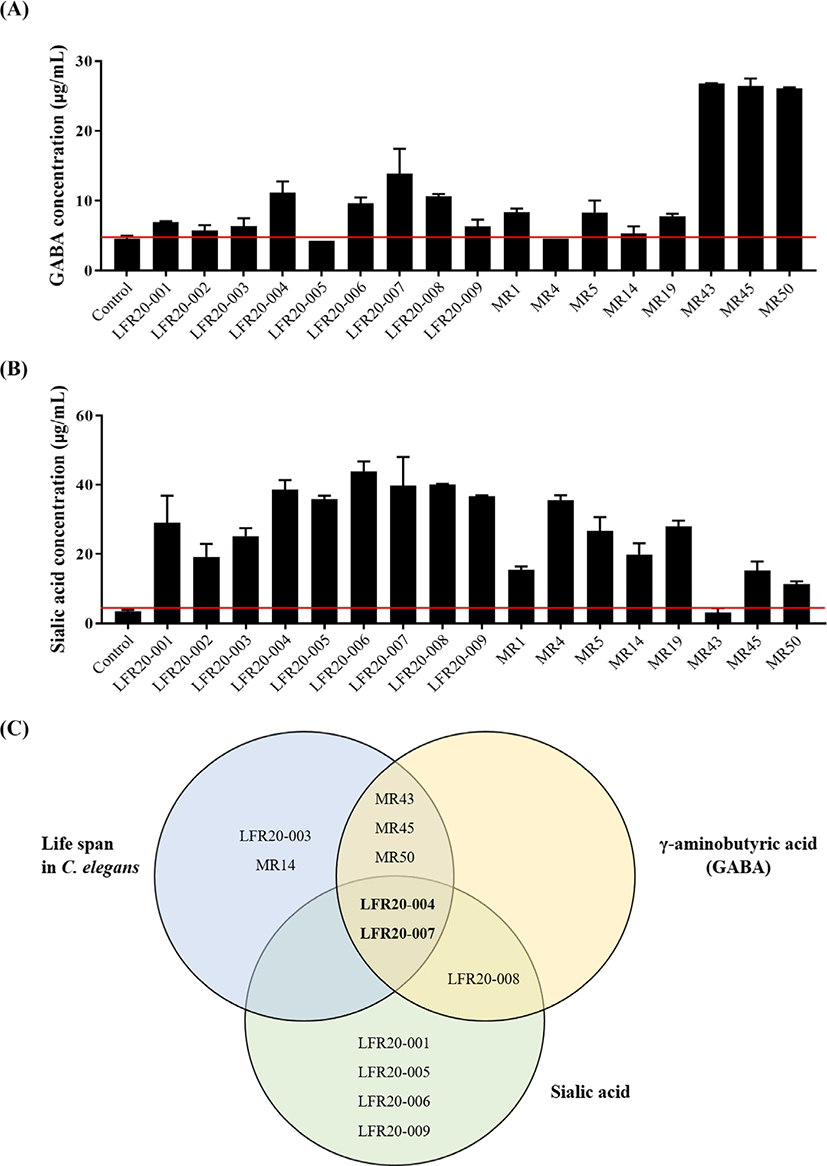
We then compared the biological activity of FAV with AV. We utilized two strains, LFR20-004 and LFR20-007, among the strains that increased the longevity in C. elegans (10 strains), GABA production (6 strains), and sialic acid content (10 strains) for the fermentation (Fig. 2C). GC-MS analysis of FAV revealed that free amino acid, short-chain fatty acid, and monosaccharide contents were changed significantly (Table 5). This profile may include metabolites derived from two strains and ingredients used by LAB or not used in AV samples.
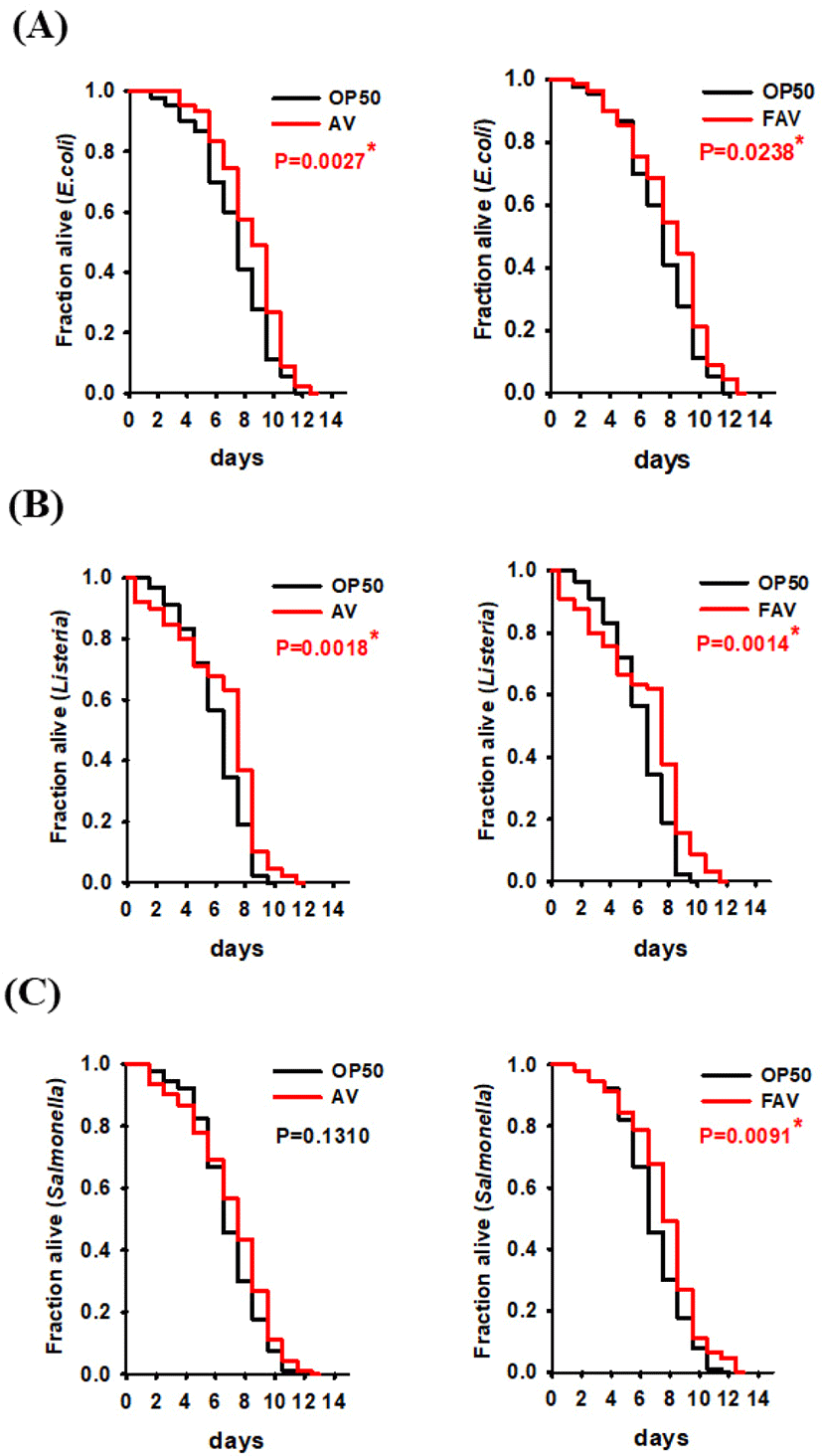
We evaluated the biological effects of FAV on the antibacterial immune response in C. elegans by assessing whether the FAV and AV with probiotics influenced the host’s survival after infection by pathogens (E. coli O157:H7, Salmonella typhi, and Listeria monocytogenes). The survival of the C. elegans conditioned with FAV or AV for 24 h was significantly higher compared with that of the control worm group fed only normal feed (non-pathogenic E. coli OP50) when exposed to two pathogens (E. coli O157:H7 and Listeria monocytogenes) (Figs. 3A and B). The worms conditioned with FAV exhibited significant resistance to infection by S. typhi, while worms conditioned with AV were not different from those fed normal feed (Fig. 3C). There have been few studies on the antibacterial efficacy of deer extracts against pathogens. To our knowledge, this study is the first to report that non-fermented deer antlers increase resistance of C. elegans to E. coli O157:H7 and Listeria monocytogenes. Dai et al. [35] reported that antler velvet extracts stimulated anti-inflammatory cytokines in splenocytes and phagocytes and play a protective role in Staphylococcus aureus-infected mice. However, fermentation products are known to have excellent antibacterial efficacy owing to the production of short-chain fatty acids such as lactic acid and play an important role in the safe storage of fermented foods [36–39]. Our results showed that the FAV increased resistance of C. elegans to pathogenic bacteria.
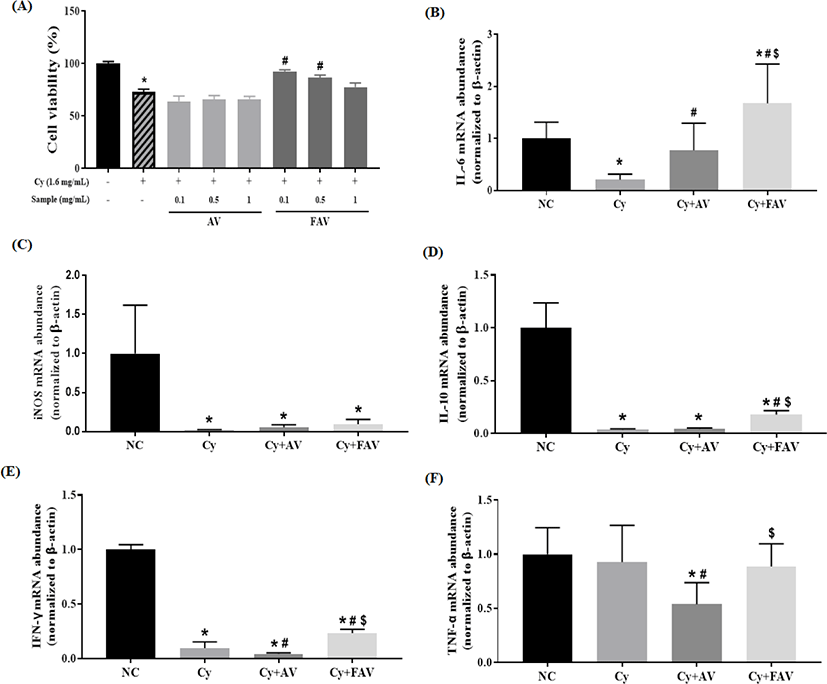
To evaluate the effects of FAV and AV on cytotoxicity of Cy, an immune-suppressing agent, to primary splenocytes, cells isolated from spleens of mice were incubated with various concentrations of AV or FAV (0, 0.1, 0.5, or 1 mg/mL) and Cy (1.6 mg/mL) for 48 h. Surprisingly, we found that FAV completely restored cell viability at 0.1 mg/mL, which was the lowest concentration. However, the viability of splenocytes treated with AV remained reduced by Cy (Fig. 4A). This finding indicated the protective effect of FAV against Cy-induced cell death and encouraged us to induce immunosuppressed mice by administering Cy (100 mg/kg). Expression of various immune modulators such as interleukin (IL)-6, IL-10, interferon (IFN)-γ, inducible nitric oxide synthase (iNOS), and tumor necrosis factor (TNF)-α by activated macrophages has been reported to play an important role in host defense against harmful factors [40]. Cytokines such as IL-6 and TNF-α exhibit essential immune functions during early stages of infection [41,42]. IFN-γ, which is a type-1 T-helper (Th1)-derived cytokine, also plays important roles in cell-mediated immune response [43]. IL-6 and IL-10 are type-2 T-helper (Th2)-derived cytokines that promote humoral immunity [44]. NO is synthesized by iNOS, an important molecule that activates macrophages to neutralize pathogenic microorganisms and tumor cells [45]. We analyzed mRNA expression of immune modulators such as IL-6, IL-10, iNOS, IFN-γ, and TNF-α in splenocytes isolated from immunosuppressed mice. We observed that FAV promoted the production of immunomodulators in spleen tissue-derived cells while AV did not (Fig. 4). In addition to primary splenocytes, it is necessary to investigate the effect of FAV on the single type of isolated immune cells, such as natural killer cells, T/B cells, neutrophils, eosinophils, and dendritic cells, to describe the biological mechanisms for the immune-enhancing activity of FAV.
The health-promoting effects of antler velvet in terms of chemical composition, bioactive components, and pharmacological effects have been reported [46]; however, there is not adequate evidence for its immune-enhancing efficacy. Zhoa et al. have suggested that peptides (3.2 kDa) extracted from antler velvet can modulate the immune system via regulation of Th1- and Th2-related cytokines [47]. Consistently, in our study, antler velvet fermented with two bacterial strains stimulated the immune system while AV did not.
The global demand for health- and immune-boosting functional foods has increased in the COVID-19 era, and studies on the development of immune-promoting substances and food processing are also increasing [48]. In this study, we demonstrated that FAV increased resistance to pathogenic bacteria in C. elegans and promoted splenocyte activation in mouse models by stimulating immune modulators. As deer antler velvet is fermented with LFR20-004 and LFR20-007, the bioactive molecules (GABA and sialic acid) as well as the immune function of deer antler velvet is expected to improve. In the future, an in-depth analysis of the metabolites from the LAB, L. rhamnosus LFR20-004 and L. sakei LFR20-007 that exhibit immune-promoting functions will be required. These LAB can be used as probiotic starters for health functional foods fermented for deer antler velvet as LAB.