
INTRODUCTION
The immune system comprises innate and adaptive immunity. It is a tightly regulated network that maintains homeostasis under normal physiological conditions [1]. Lymphocytes and macrophages play vital roles in both types of immunity [2,3]. Natural killer (NK) cells are key components of innate immunity and guard the host against tumors and viral infections. T and B lymphocytes are primary effectors in adaptive immunity [4]. Activated lymphocytes and macrophages produce inflammatory mediators such as nitric oxide (NO), tumor necrosis factor (TNF)-α, interferon (IFN)-γ, and interleukin (IL)-12 [5–7].
Probiotics are living microorganisms that provide various health benefits to the host, and lactic acid bacteria (LAB) such as Lactobacillus and Bifidobacterium are known as common probiotic bacterial species [8,9]. Particularly, the immunomodulatory efficacy of probiotics has aroused much research interest in recent years [10,11]. However, their effects on the immune system widely vary even among strains within the same species. Hence, characterizing the effects of specific strains and determining their optimal doses and combinations are necessary to validate the health claims [12]. Previous studies have demonstrated that LAB-induced immunomodulatory responses are mediated through pattern recognition receptors (PRRs) expressed on immune cells, such as toll-like receptors (TLRs) [13–15]. However, even in the same species of LAB, the PRRs mainly involved are different depending on the strain, and the immunomodulatory effect may appear in various ways, such as immunostimulation and immunoregulation [16].
Cyclophosphamide (CTX) is an antineoplastic and immunosuppressive drug administered for various cancers and autoimmune diseases [17,18]. However, it produces activated metabolites that interfere with DNA replication and damage mitochondrial and lysosomal membranes, which can injure normal cells as well as cancer cells [19]. The CTX-induced cytotoxicity may generate various side effects such as hematological malignancies, infections, and bone marrow depression [20]. Hence, researchers have investigated immunoregulatory agents such as functional foods and dietary supplements that mitigate the side effects of chemotherapeutic agents [21].
Bifidobacterium animalis ssp. lactis HY8002 (HY8002) is a probiotic strain confirmed to improve intestinal integrity disturbed by antibiotics [22] and reduce airway hypersensitivity and inflammatory response caused by fine dust [23]. Nevertheless, its effect and mechanism on the systemic immune response are unclear. In addition, Lactobacillus plantarum HY7717 (HY7717) was selected as a new potential immunostimulatory candidate. Although there have been studies on the immunostimulatory effects of individual strains of probiotics, there is insufficient evidence on the effects of the combination of Bifidobacterium and Lactobacillus strains on systemic immunity and immunosuppression improvement [24–27]. Therefore, we explored possible synergy of efficacy between verified probiotic Bifidobacterium strain HY8002 and the newly selected Lactobacillus strain HY7717 in mouse primary splenocytes and CTX-induced immunosuppressed mice and aimed to elucidate their molecular mechanisms.
MATERIALS AND METHODS
Roswell Park Memorial Institute (RPMI) 1640 medium, Dulbecco’s modified Eagle minimal essential medium (DMEM), antibiotic-antimycotic, and fetal bovine serum (FBS) were obtained from Gibco (Grand Island, NY, USA). Griess reagent, levamisole hydrochloride (LH), CTX, concanavalin A (Con A), and lipopolysaccharide (LPS) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Enzyme-linked immunosorbent assay (ELISA) kits for IFN-γ, IL-12, and TNF-α were acquired from BD Biosciences (San Diego, CA, USA). ELISA kits for IgA and IgG were procured from Abcam (Cambridge, MA, USA) and Invitrogen (Carlsbad, CA, USA), respectively. Anti-mouse TLR2 monoclonal antibody (mAb) (clone C9A12. No. mabg-mtlr2) and anti-mouse TLR4 mAb (clone MTS510. No. 117617) were obtained from InvivoGen (San Diego, CA, USA) and BioLegend (San Diego, CA, USA), respectively.
LAB strains including B. animalis ssp. lactis HY8002 and L. plantarum HY7717 were stored in the seed culture library of hy (Yongin, Korea). Bifidobacterium strains and Lactobacillus strains were anaerobically cultured in blood glucose liver (BL) broth or de Man Rogosa and Sharpe (MRS) broth at 37°C for 18 h. The cultured LAB strains were centrifuged at 2,000×g for 15 min and then resuspended in PBS (phosphate buffered saline) for subsequent experiments.
NO release was indirectly detected by measuring stable NO catabolites in cell culture media via the Griess reaction. RAW 264.7 macrophages were plated into 96-well plates at 1.0 × 104 cells/well and treated with LAB strains (1.0 × 105 cells/well) for 24 h. The cell culture medium was mixed with an equal amount of Griess reagent and left in the dark at room temperature for 15 min. Absorbance was measured at 540 nm and the NO quantity was determined by interpolation of a sodium nitrite standard curve.
Splenocytes were isolated from aseptically collected mouse spleens through a 40 μm cell strainer, and then plated at 1.0 × 106 cells/well in RPMI 1640 medium in a 24-well plate. Each well was treated with 1.0 × 107 CFU/mL of HY8002, HY7717, or a 1:1 mixture of HY8002 and HY7717, and then cultured at 37°C under 5% CO2 conditions for 24 to 48 h. Cytokine levels in the culture medium were determined with ELISA kits according to the manufacturer’s instructions.
RNA was extracted using an Easy-BLUETM Total Extraction kit (iNtRON Biotechnology, Seongnam, Korea). The cDNA was synthesized with an Omniscript® Reverse Transcription kit (Qiagen, Hilden, Germany) on a thermal cycler (Bio-Rad Laboratories, Hercules, CA, USA). The mRNA levels were measured using a TaqMan Universal PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and a QuantStudio 6 Real-Time PCR System (Applied Biosystems). The TLR2 (Mm00442346_m1), TLR4 (Mm00445273_m1), and GAPDH (Mm99999915_g1) transcripts were quantified with gene-specific primers.
Male Balb/c mice (7 weeks; 20–22 g) were obtained from Dooyeol Biotech (Seoul, Korea). The animal facility was maintained at 20°C–22°C, 40–60% relative humidity (RH), and 12 h light/12 h dark cycle. The mice had ad libitum access to an autoclaved standard laboratory diet and water. All experimental procedures were approved by the Ethics Review Committee of the R&D Center of hy Co. Ltd., Korea (IACUC approval number, AEC-2021-0009-Y). After 1 week adaptation, the mice were divided into six groups (n = 5): normal mice (NOR), CTX, LH (20 mg/kg/day), HY8002 (1 × 109 CFU/kg/day), HY7717 (1.0 × 109 CFU/kg/day), and HY8002+HY7717 (1:1 ratio; 1.0 × 109 CFU/kg/day). LH, HY8002, and HY7717 were intragastrically administered in sterile saline once daily for 8 day. All mice except those in the NOR group were intraperitoneally injected with 150 mg/kg/day CTX on day 8. At day 10, whole blood was collected in anticoagulant tubes, the plasma was isolated, and the spleens were excised and weighed.
Splenic NK cells were isolated by Magnetic-activated cell sorting (MACS) Cell separate kit with CD49b (DX5) MicroBeads (Miltenyi Biotec Technology, Bergisch Gladbach, Germany), placed in a 96-well round-bottom plate at a density of 1.0 × 105 cells/well and co-cultured with an equal density of YAC-1 cells for 16 h. Lactate dehydrogenase (LDH), as an indicator of NK cell cytotoxicity against YAC-1, was measured using a CytoTox® 96 non-radioactive cytotoxicity assay kit (Promega, Madison, WI, USA).
Splenocytes were prepared as previously described, plated into 96-well plates at a density of 2.0 × 105 cells/well, and subjected either to Con A (4 μg/mL) or LPS (1 μg/mL). After 48 h, cell counting kit-8 (CCK-8) solution was added and absorbances were measured at 450 nm.
Blood counts were performed using an auto hematology analyzer (BC-5000 Vet; Mindray, Shenzhen, China).
Plasma IgA, IgG, and IL-12 levels were measured using ELISA kits according to the manufacturer’s instructions.
Data are represented as means ± SD for ≥ 3 independent experiments. Significant differences were identified by one-way analysis of variance (ANOVA) followed by the Newman–Keuls test (p < 0.05). An unpaired t-test was used to compare means between two independent groups. All statistical analyses were performed using GraphPad Prism v5 (San Diego, CA, USA).
RESULTS
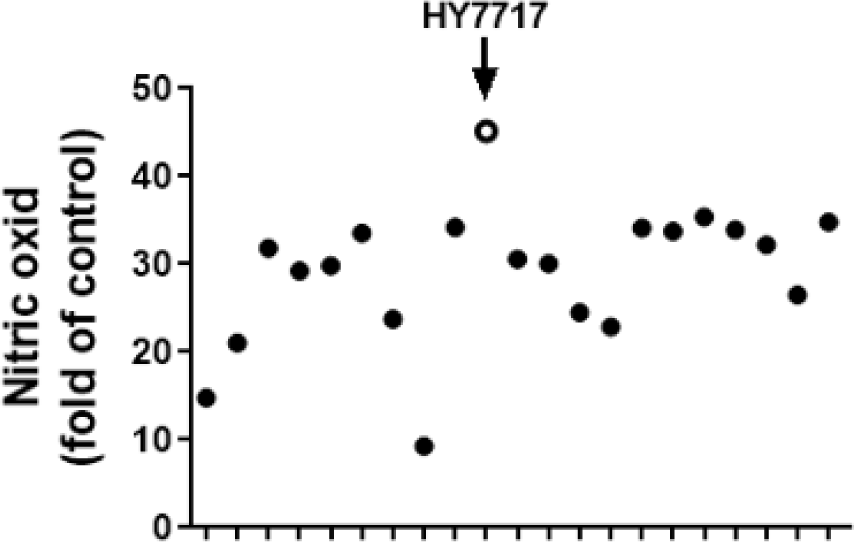
We tested the effects of 252 LAB strains on NO inhibition in RAW 264.7 macrophages. Of these, 21 demonstrated anti-inflammatory efficacy (data not shown) and their immunostimulatory function was evaluated. We measured NO concentrations in culture media to determine whether the 21 LAB strains activate RAW 264.7 (Fig. 1). Eleven of the strains increased the relative NO rates by > 30-fold. Of these, HY7717 showed the largest NO rate increase (45.2-fold).
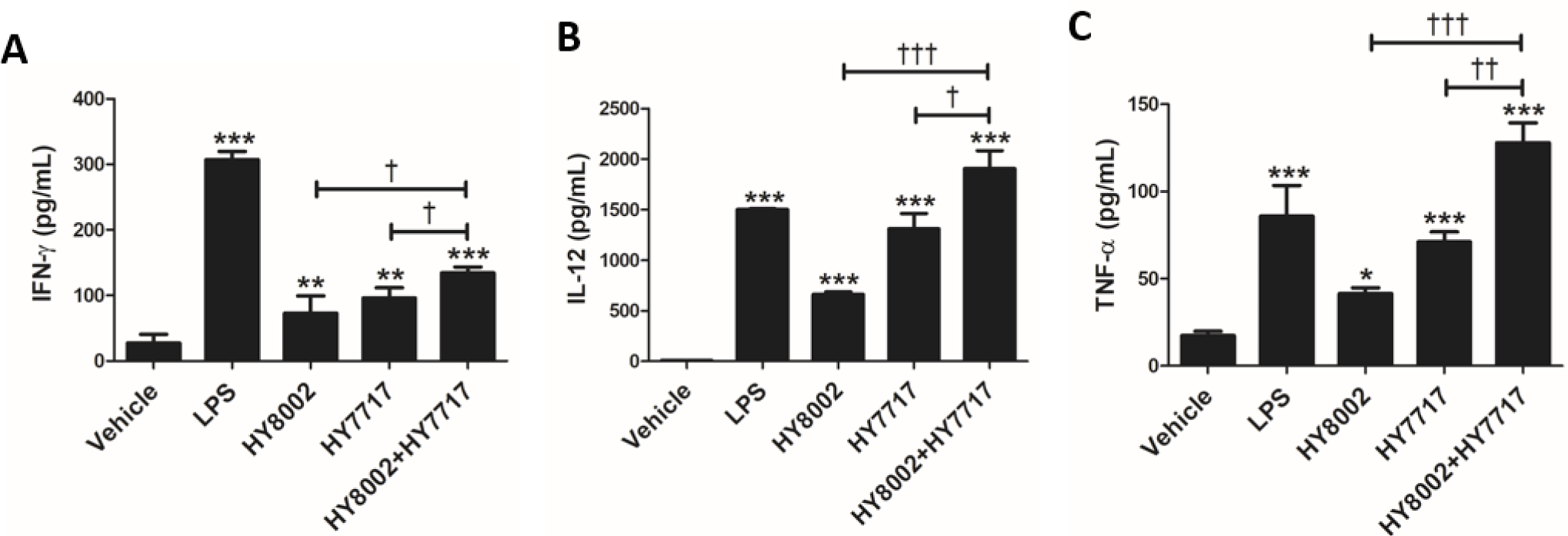
We subjected splenocytes to single and combined HY8002 and HY7717 treatments and measured the IFN-γ, IL-12, and TNF-α levels using ELISA (Fig. 2). The cytokine levels in the splenocytes treated with all LAB strains were significantly higher than those in the vehicle control. Cytokine secretion was significantly higher in response to all combination treatments than each standalone treatment.
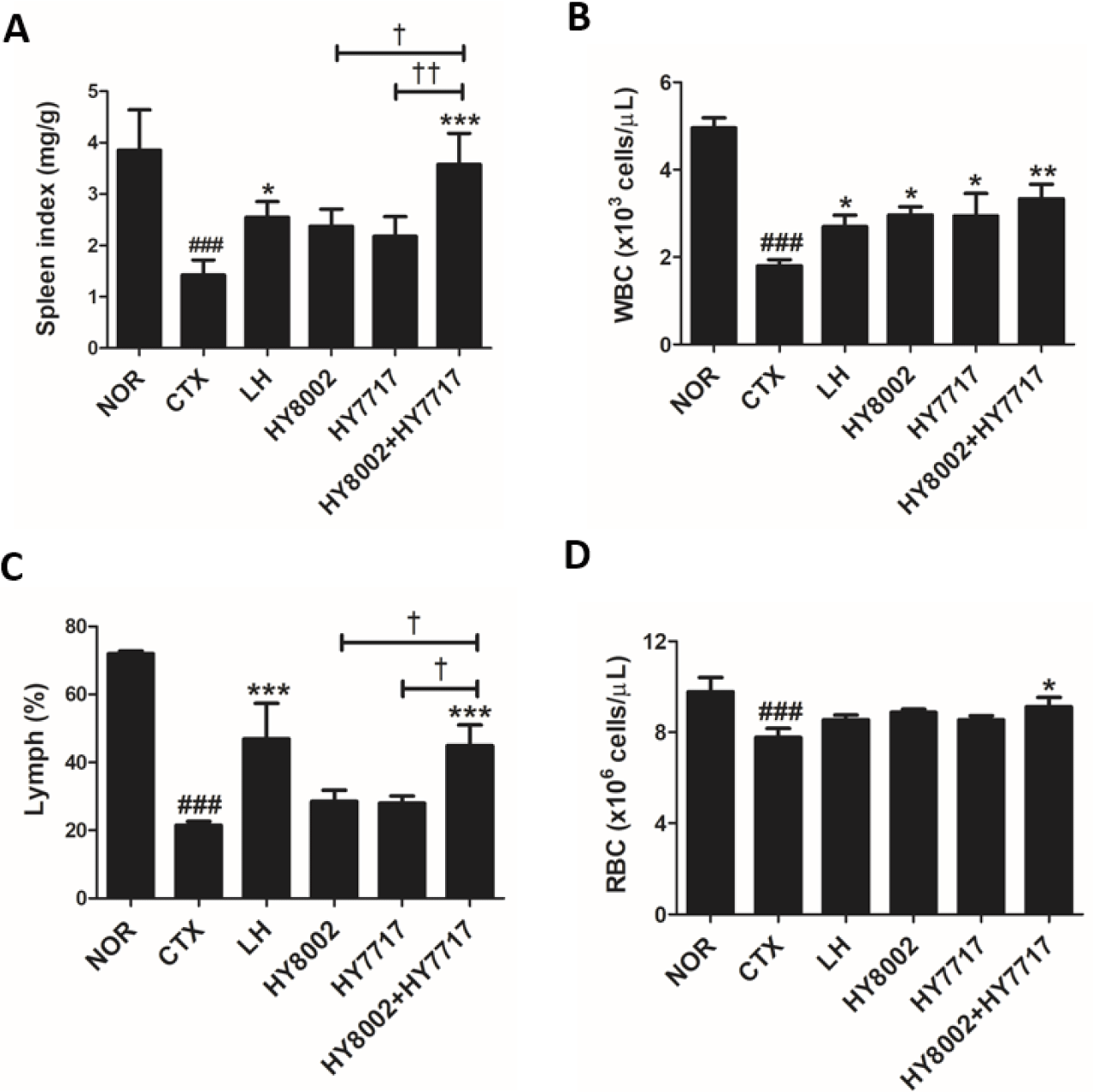
HY8002, HY7717, or a mixture of both was administered to CTX-immunosuppressed mice to assess the immunostimulatory efficacy of LAB strains under physiological conditions. The CTX treatment markedly reduced the spleen index (Fig. 3A). The spleen indices were significantly higher in the LH and HY8002+HY7717 groups than the CTX group. Oral administration of each strain alone non-significantly increased the spleen indices. We also measured the complete blood count (CBC) parameters in each group (Figs. 3B–3D). Compared with the NOR group, the white blood cells (WBC) and red blood cells (RBC) counts and Lymph% in the CTX group were significantly reduced by 36.4%, 79.4%, and 29.9%, respectively. LH administration significantly increased the relative WBC count and Lymph% but not the RBC count. The HY8002, HY7717, and HY8002+HY7717 groups presented with significantly higher WBC counts than the CTX group. The HY8002+HY7717 group presented with significantly higher Lymph% and RBC counts than the CTX group. However, the HY8002 and HY7717 groups presented with non-significantly higher Lymph% and RBC counts than the CTX group.
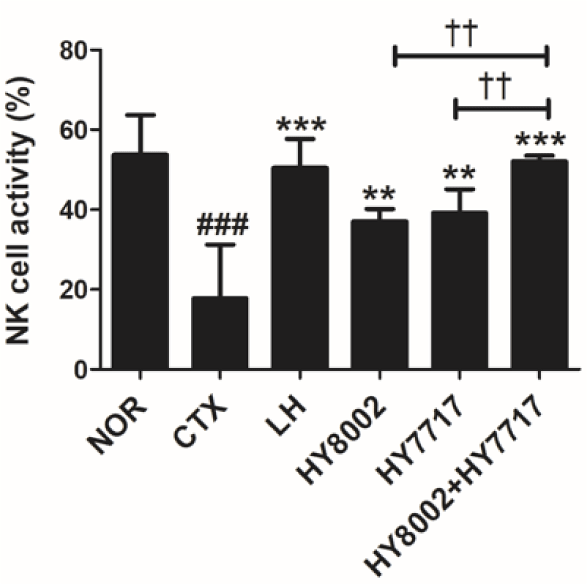
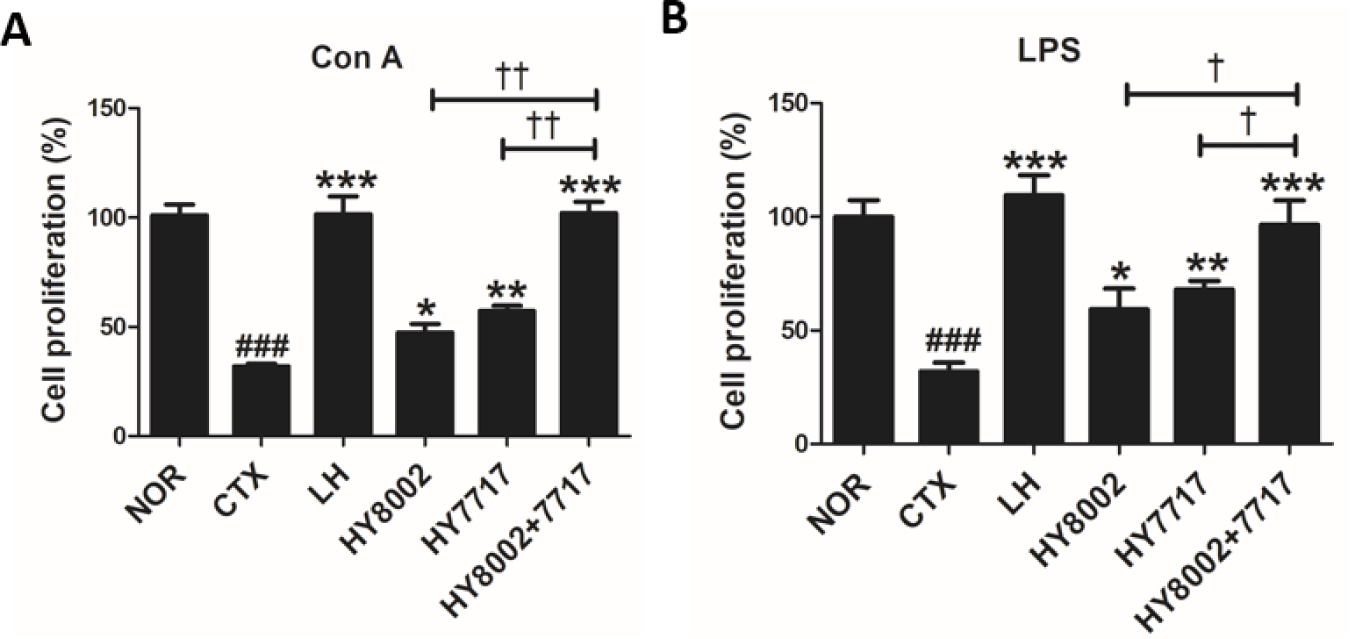
NK cell activity and T and B cell proliferation were assessed for the spleens of all treatment groups. The CTX treatment significantly decreased NK cell cytotoxicity against its YAC-1 target cell (Fig. 4). However, the LH treatment restored NK cell activity. All LAB treatments significantly increased NK cell activity. However, the combination treatments were significantly more efficacious than the single treatments. Splenic lymphocytes were incubated with Con A and LPS to evaluate T and B lymphocyte proliferation, respectively (Fig. 5). LH reversed CTX-induced inhibition of T and B lymphocyte proliferation. Standalone HY8002 and HY7717 and their combination significantly increased T and B lymphocyte proliferation compared with the CTX group. Moreover, combined LAB strain administration significantly enhanced T and B lymphocyte proliferation compared with each standalone LAB treatment.
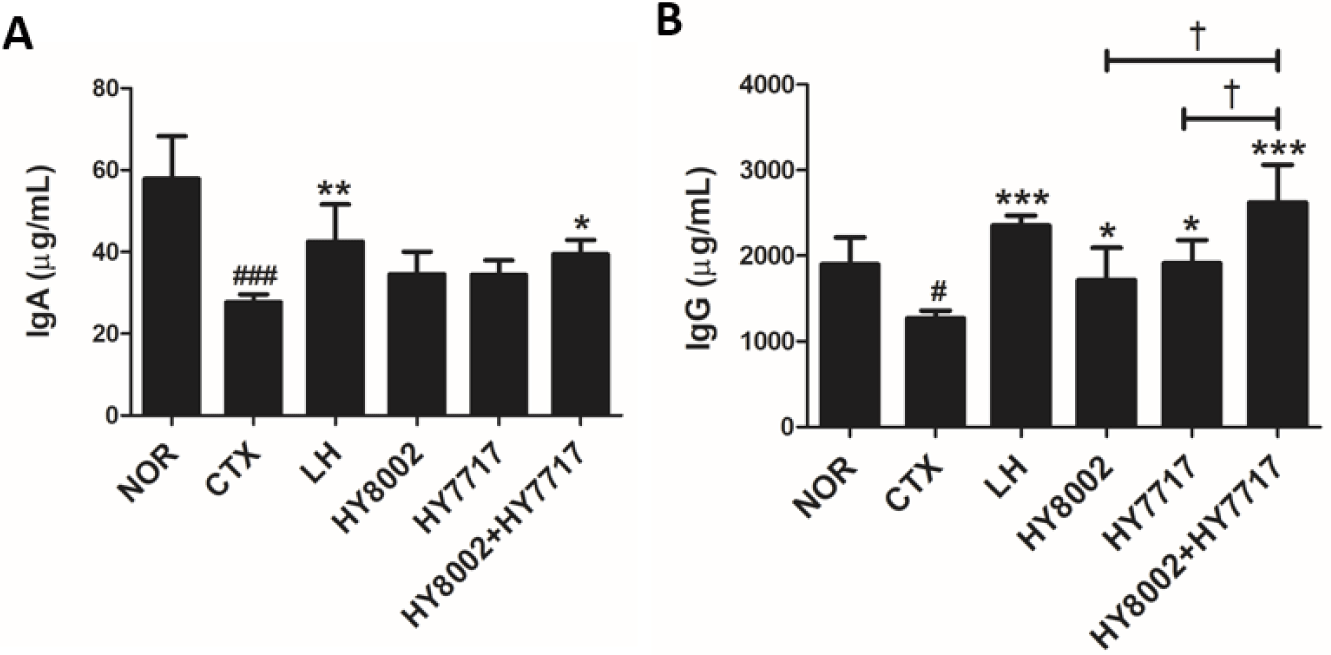
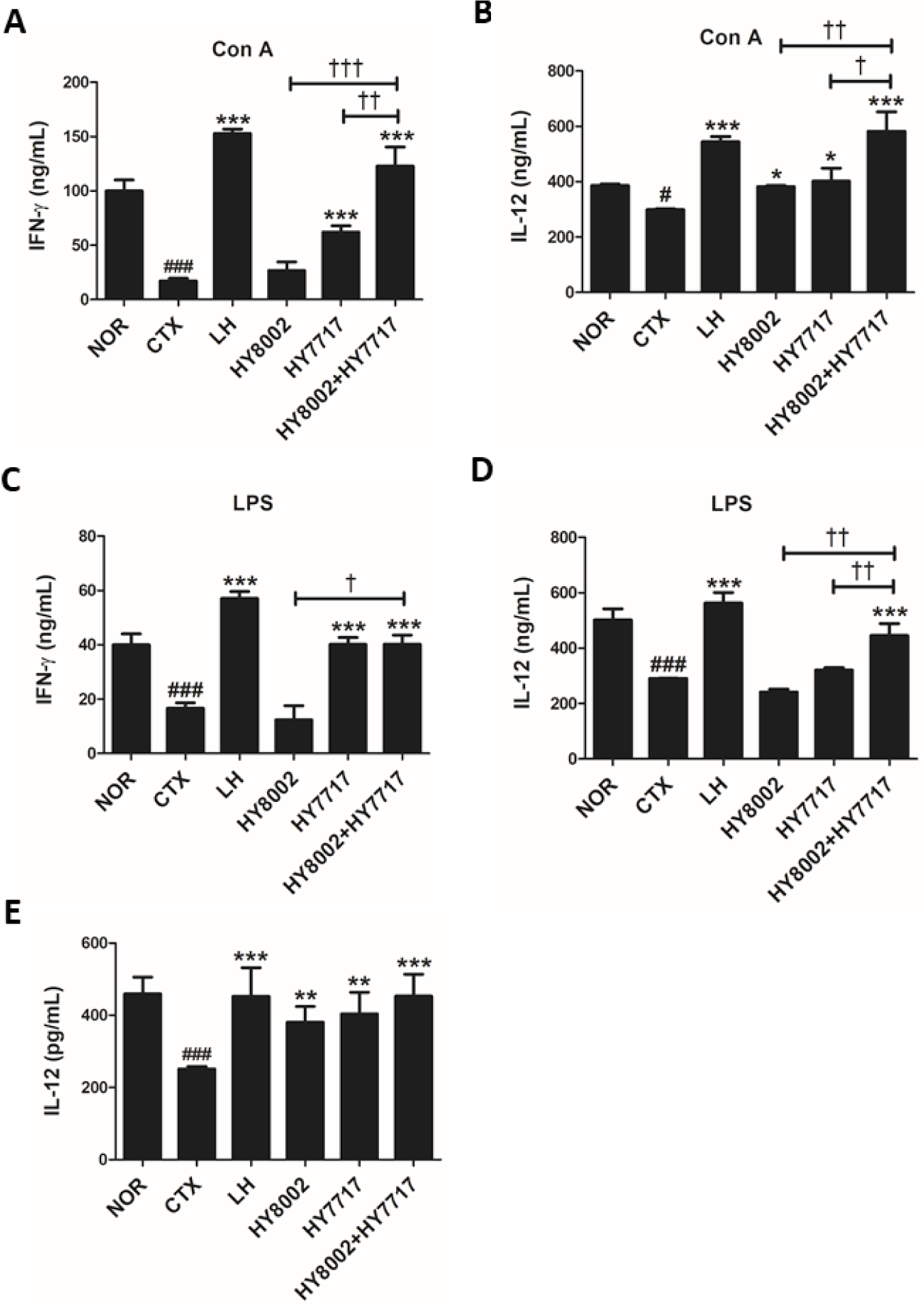
We quantified the relative plasma IgA and IgG levels (Fig. 6). Oral LH administration significantly increased IgA and IgG reduced by CTX treatment. No standalone LAB strain significantly altered the IgA levels whereas the combinations did significantly raise them. All LAB treatments significantly increased the IgG levels compared with the CTX group. However, the combination treatments significantly and more effectively increased the IgG levels than the standalone treatments. We measured IFN-γ and IL-12 secretion in cultured Con A- or LPS-stimulated splenic lymphocytes (Figs. 7A–7D). The LH and HY8002+HY7717 treatments increased the cytokine levels reduced by the CTX treatments in the Con A-activated T lymphocytes and the LPS-activated B lymphocytes. The cytokine levels in the Con A-activated T lymphocytes of HY8002+HY7717 group were significantly higher than those of standalone LAB groups. The IFN-γ levels in the LPS-activated B lymphocytes of HY7717 or HY8002+HY7717 groups were significantly higher than those of CTX group. Only HY8002+HY7717 administration significantly increased the relative IL-12 levels in the LPS-activated B lymphocytes. The HY8002, HY7717, HY8002+HY7717, and LH treatments recovered the plasma IL-12 levels reduced by CTX treatment (Fig. 7E).
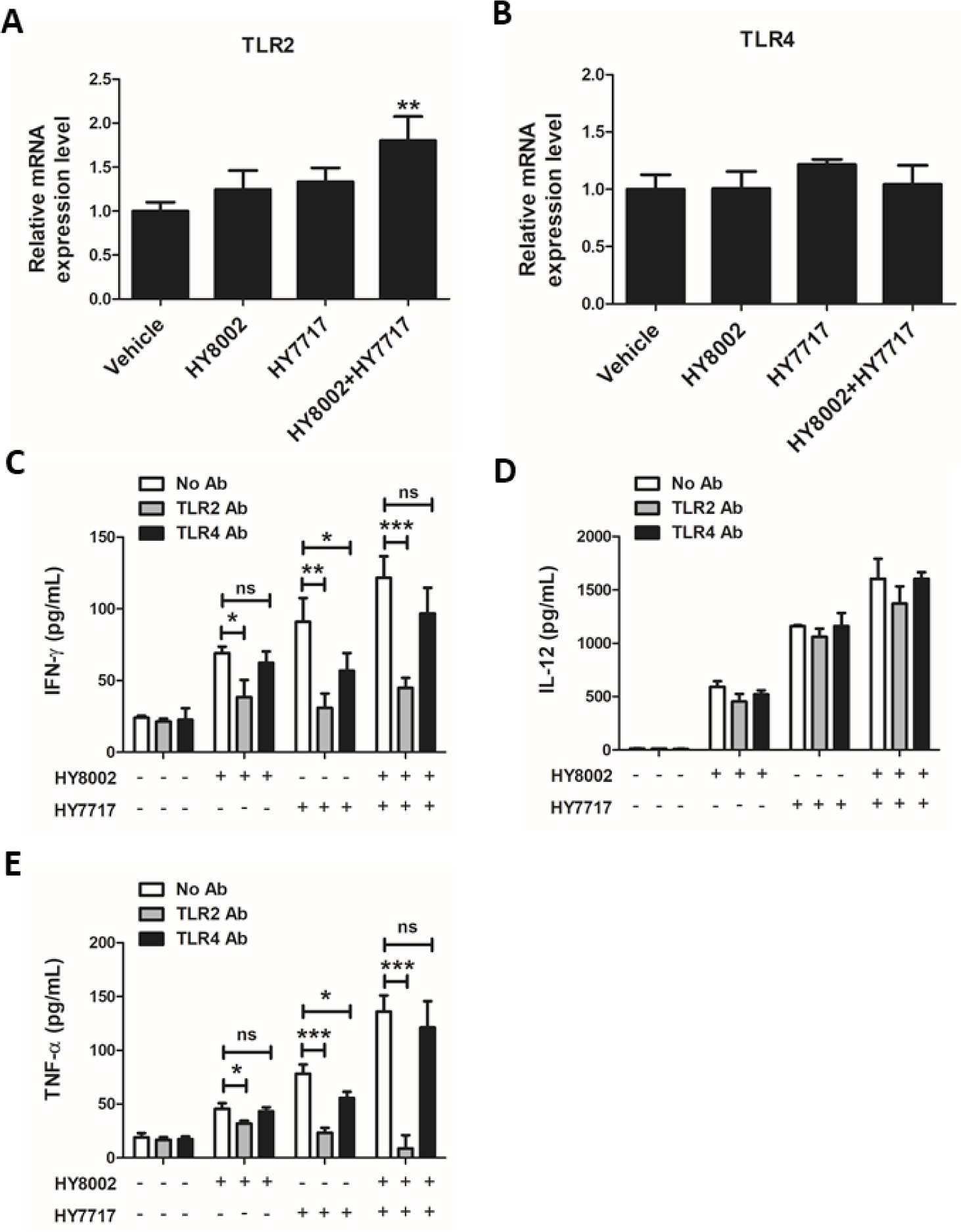
TLR2/4 mRNA expression levels in splenocytes treated with single and combined HY8002 and HY7717 were measured to determine whether the immunostimulant efficacy of these LAB is associated with TLRs (Figs. 8A and 8B). Each standalone LAB strain slightly upregulated TLR2 mRNA but the combination treatment significantly upregulated it. The HY7717 treatment non-significantly upregulated TLR4 mRNA. After pretreatment with anti-TLR2 or anti-TLR4 antibodies, the IFN-γ, IL-12, and TNF-α levels were measured in the culture media of splenocytes subjected to HY8002, HY7717, or their combination (Figs. 8C–8E). Anti-TLR2 or anti-TLR4 antibody alone had no significant inhibitory effect on the cytokine levels. Splenocytes blocked with anti-TLR2 antibody secreted significantly less IFN-γ and TNF-α than those also treated with LAB. Anti-TLR4 antibody blocked IFN-γ and TNF-α production only in the HY7717-treated splenocytes. However, neither antibody had any significant effect on IL-12 secretion.
DISCUSSION
There has been growing interest in the immunomodulatory potential of probiotics in the prevention and treatment of various complex symptoms [28]. Several LAB strains demonstrated health benefits by enhancing the immune system [29–31]. Certain LAB strains regulate T cell effector functions, enhance humoral immunity, and activate lymphocytes and macrophages [32]. Our previous work showed that the number of IFN-γ-secreting T cells was increased by co-culturing them with Bifidobacterium animalis ssp. lactics HY8002-treated dendritic cells [33]. Nevertheless, the immunostimulatory properties of HY8002 have not been thoroughly investigated. Here, we discovered that Lactobacillus plantarum HY7717 is a potential immunomodulatory agent and evaluated its immunostimulatory efficacy alone and in combination with HY8002 both ex vivo and in vivo.
Spleen is a significant organ in terms of understanding immune cell subpopulations and their immune responses to antigens. Splenocytes include T and B cells, macrophages, and natural killer (NK) cells [34]. Their immune responses include antigen presentation, T lymphocyte activation, and B lymphocyte differentiation [35]. Cytokines control immune cell proliferation and activity. Proinflammatory cytokines such as IFN-γ, IL-12, and TNF-α augment inflammatory responses [12]. IFN-γ is produced mainly by T and NK cells and activates macrophages, neutrophils, and other NK cells [36]. IL-12 is a heterodimeric protein released from antigen-presenting cells (APCs) such as dendritic cells, macrophages, and B cells and it activates NK and T cells [37]. TNF-α is produced mainly by activated macrophages, T cells, and NK cells and promotes inflammatory activity in macrophages [38]. Here, HY8002 and HY7717 stimulated IFN-γ, IL-12, and TNF-α production in splenocytes. Moreover, the combination of HY8002 and HY7717 effectively increased cytokine production.
The mouse CTX-induced immunosuppression model is widely used to investigate immunomodulatory activity. CTX inhibits the immune response by inactivating immune cells and decreasing proinflammatory cytokine production [39]. Here, we investigated the immunomodulatory properties of HY8002 and HY7717 in a mouse CTX-induced immunosuppression model. LH is immunostimulatory, has several adverse effects, and was used as the positive control [40,41]. As the spleen is a major organ in the immune system, its size is a vital indicator of health and disease [42]. Moreover, changes in the lymphocyte count are used to diagnose immune-mediated disease [43]. Recent studies showed that red blood cells (RBCs) modulate immune cell activity and maturation [44,45]. In the present study, we confirmed that CTX induced immunosuppression by reducing spleen and hematological indices and verified that administration of a combination of HY8002 and HY7717 increased these indices and counteracted the negative effects of CTX.
NK cells mediate antitumor and antiviral immune responses [46], by recognizing infected cells in the absence of major histocompatibility complex (MHC) and enable rapid immune responses. T and B lymphocyte proliferation is vital in adaptive immune system activation, and their proliferation in response to mitogens has been widely applied to evaluate the sensitivity of these cells [47]. Here, HY8002 and HY7717 effectively enhanced NK cell activity and T and B cell proliferation in CTX-treated mice.
Immunoglobulin A (IgA) is the first-line defense against infection while IgG is the major antibody in secondary responses. Proinflammatory cytokines play crucial roles in inflammation as a host defense mechanism. Inflammatory responses are beneficial when cytokines are secreted at appropriate level [36]. In the present study, a combination of HY8002 and HY7717 upregulated IgA and IgG in CTX-treated mice. It also increased IFN-γ and IL-12 secretion in Con A-activated T cells more than either treatment alone. The foregoing combination also increased IFN-γ and IL-12 production by LPS-activated B cells. Plasma IL-12 levels were higher in response to this combination than either treatment alone. Therefore, administration of HY8002 plus HY7717 resisted immunosuppression in vivo.
TLRs modulate both innate and adaptive immunity. Certain TLRs are co-stimulatory and increase proliferation and cytokine production in T cells stimulated by their respective ligands [48]. Another study demonstrated that TLR signaling is implicated in cytokine production by B cells, antigen presentation, Ig class switching, and B cell survival [49]. The bioactive factors and microbe-associated molecular patterns (MAMPs) produced by different LAB strains activate TLR2 [14,50] Administration of certain LAB strains increased the relative numbers of TLR4+ and TLR2+ cells produced by mice [51]. Here, we investigated whether TLRs mediate HY8002 and HY7717 immune responses. The TLR2 mRNA expression levels increased in healthy splenocytes in response to a combination of HY8002 plus HY7717. By contrast, none of the treatments significantly modulated the TLR4 mRNA expression levels. We demonstrated that anti-TLR2 antibody blocked LAB-induced IFN-γ and TNF-α secretion but had no inhibitory effect on IL-12. Thus, the immune responses of HY8002 and HY7717 may be associated with PRRs such as other TLRs or NOD-like receptors (NLRs) [52]. TLR2 is a major receptor of both HY8002 and HY7717 and promotes immune activity in splenocytes. Weaning stress in piglets can suppress systemic immune responses, making piglets vulnerable to infectious diseases, including diarrhea. Jing Wang et al. reported that the supply of Lactobacillus plantarum improves intestinal health in weaned pigs by increasing the expression of host defense peptide (HDP) through TLR2 signaling [53]. Since HY7717 and HY8002 have the effect of improving systemic immunosuppression, they are expected to be effective in improving the immune load caused by weaning stress in piglets [54]. However, it may not be the sole receptor involved in this process. Therefore, further studies are needed on factors other than TLRs that may influence HY8002 and HY7717-mediated immune responses.
In this study, the combination of Bifidobacterium animalis ssp. lactics HY8002 and Lactobacillus plantarum HY7717 was more efficacious than either treatment alone at stimulating cytokine release in mouse primary splenocytes. HY8002 plus HY7717 effectively rehabilitated the splenic and hematological indices, NK cell activity, T and B cell proliferation, and immunoglobulin and cytokine production in a mouse CTX-induced immunosuppression model. The immunostimulatory effects of HY8002 and HY7717 were closely associated with TLR2. Hence, the combination of HY8002 and HY7717 effectively enhances immune function, suggesting the possibility of this combination as a probiotics for immune enhancement. Collectively, the combination of the two probiotics would be considered a potential candidate to application of functional dairy foods.