Enterococcus faecium (E. faecium) is recognized for its biological properties, which include the ability to inhibit harmful bacteria, good adhesion to the intestinal mucosa, and resistance to bile salts, stomach acid, and heat [1]. Because of these properties, it is widely used as probiotics for humans and animals. Previous studies have shown that adding E. faecium as a probiotics to weaning piglet feed improved growth and feed conversion [2]. Enterococcus is dominant microbiota of Doenjang, a traditional Korean fermented food made from soybeans [3].
In this study, we isolated E. faecium strain AJ_C_05 from a homemade soybean paste, Doenjang, collected from a local market in Cheonan-si, Chungcheongnam-do, Republic of Korea. E. faecium strain AJ_C_05 was aerobically cultured in Enterococcosel broth (MBcell) at 37°C for 24 hours. The genomic DNA of the AJ_C_05 strain was extracted using the TaKaRa MiniBEST Bacterial Genomic DNA Extraction Kit Ver. 3.0 (TaKaRa) following the manufacturer’s instructions. The complete genome of E. faecium AJ_C_05 was sequenced using the Oxford Nanopore Technologies MinION platform. Briefly, a library was prepared using an Oxford Nanopore Ligation Sequencing Kit (SQK-LSK109) (Oxford Nanopore), and the sequencing process was performed on a MinION sequencing device equipped with a MinION flow cell (R9.4.1) (Oxford Nanopore). A total of 322,486,606 base pairs (bp) with 91,650 reads were obtained, and genome assembly was performed de novo using the Canu assembler (v. 1.8) and Flye assembler (v. 2.9.2) with ‘nano-corr’ options. The Homopolish polisher (v. 0.4.1) was utilized to correct errors in the assembled draft genome. Quality Assessment Tool for Genome Assemblies (QUAST) (v. 5.0.2) and Benchmarking Universal Single-Copy Orthologs (BUSCO) (v. 5.4.6) were used to quantitatively evaluate the completeness of the genome assembly. For genome functional annotation, the Rapid Annotation using Subsystem Technology (RAST) (v. 2.0) and EggNOG-mapper (v. 2.0) were utilized [4,5]. In addition, ResFinder (v.4.4.0) and BLASTn method with the Virulence Factor Database (VFDB) were used to identify antibiotic resistance genes and virulence factors, respectively.
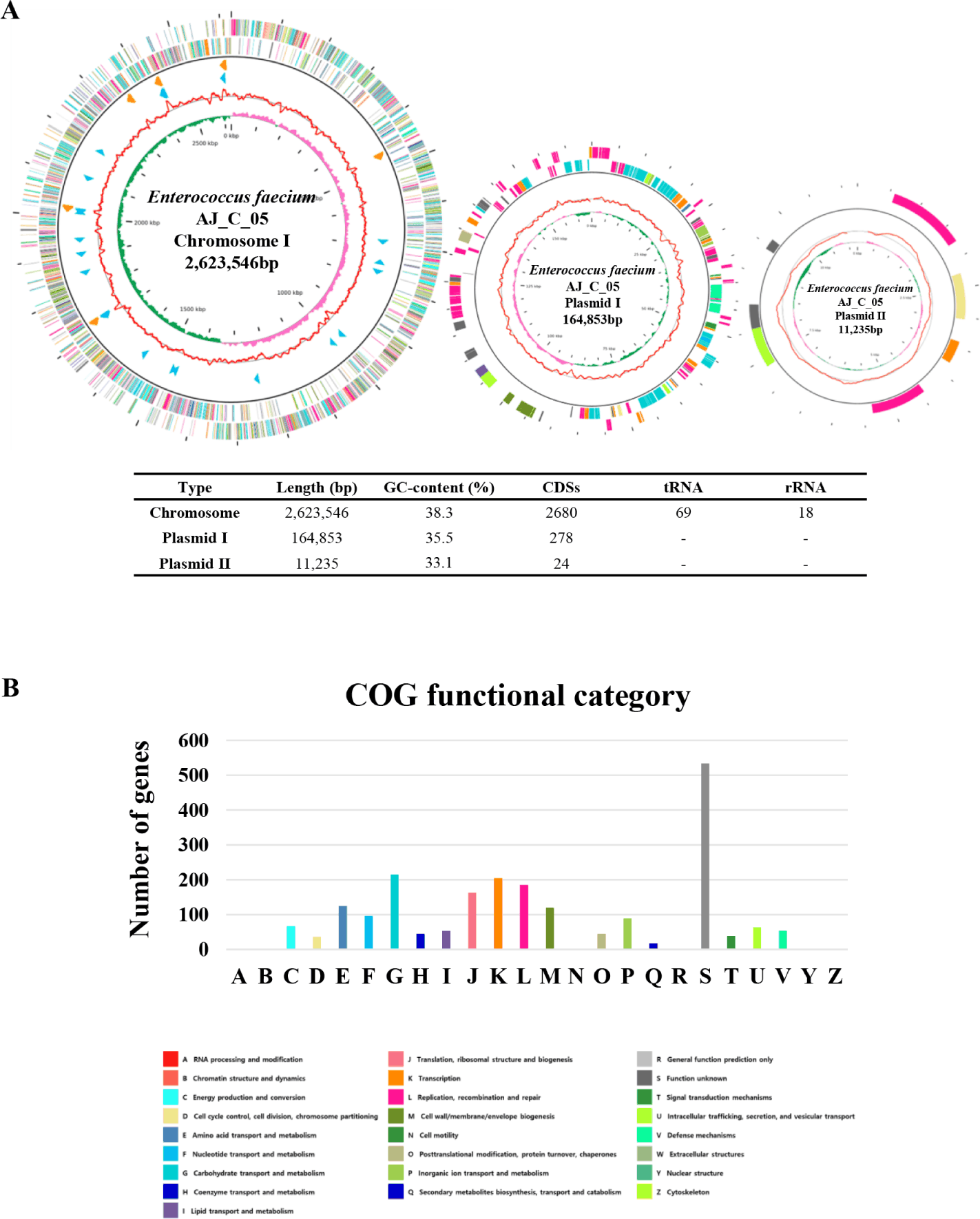
The E. faecium strain AJ_C_05 consists of the one circular chromosome and two plasmids. The chromosome genome comprises 2,623,546 bp with a guanine + cytosine (G+C) content of 38.3%, 2,680 coding sequences, 69 tRNAs, and 18 rRNAs. One plasmid is 164,853 bp long with a G+C content of 35.5% and the other is 11,235 bp long with a G+C content of 33.1% (Table 1, Fig. 1A). We compared the whole genome of E. faecium strain AJ_C_05 with that of E. faecium T110 (NCBI accession number: PRJNA207757), a strain known for its probiotic properties isolated from human microbiome in Japan [6]. E. faecium T110 has a chromosome size of 2,693,877 bp, a G+C content of 38.4%, and 2,639 protein-coding genes. The two strains have similar chromosome size, G+C content, and protein-coding genes, and an ANI value of 97.80%, indicating a high degree of similarity. Excluding an ‘unknown function’, the most COGs categories of E. faecium AJ_C_05 were ‘carbohydrate transport and metabolism [G]’, ‘Transcription [K]’, ‘Replication, recombination and repair [L]’, ‘Translation, ribosomal structure and biogenesis [J]’, ‘Amino acid transport and metabolism [E]’ and ‘Cell wall/membrane/envelope biogenesis [M]’ (Fig. 1B).
E. faecium strain AJ_C_05 possesses genes for enzyme determinants, such as cellulases (EC 3.2.1.4), proteases (EC 3.4.21.112), and xylanase (EC 3.2.1.8). Xylanase is an enzyme that specifically targets xylans, which are key carbohydrate components in the cell walls of grains [7]. Cellulase can hydrolyze non-starch polysaccharides (NSP) and destroy cell walls [8]. Protease contributes to reducing nitrogen emissions into the environment by optimizing protein utilization [9]. In addition, E. faecium strain AJ_C_05 contains the genes for enzymes such as beta-glucosidase (EC 3.2.1.21), chitinase (EC 3.2.1.122), and alpha-galactosidase (EC 3.2.1.22) that can break down carbohydrates.
The complete genome for E. faecium strain AJ_C_05 reveals that the antibiotic resistance gene aac (6’)-Ii is located on the chromosome. However, its transfer to other bacteria is hindered because of its presence on the chromosome. The pilF, encoding PilA-type pilus structure, was detected on the plasmid I. Despite being identified as a virulence factor, it may also contribute to bacterial colonization on cell surface structures, potentially benefiting probiotic gut colonization [10]. The characteristics of E. faecium strain AJ_C_05 suggests its potential for use as probiotic feed additives. This study provides valuable insights into the functionality of the newly observed E. faecium strain AJ_C_05 isolated from Doenjang, which has the potential to aid digestion when used as a bacterial feed additive for livestock.