
INTRODUCTION
pH value in the meat of pigs is associated with meat color, drip loss and moisture holding capacity; as pH increases, drip loss and cooking loss decrease, but moisture holding capacity increases, thereby affecting meat quality [1]. Meat pH value in pigs has been a subject of extensive research [2–5] it is well known that changes in pH after slaughter are of great importance in protein denaturation and drip loss in post mortem changes in muscle [6].
Investigating genetic characteristics related to meat pH value in domestic pigs is currently a priority because it involves basic stages in genomic selection using accumulated genomic information. If a reference population that increases selection accuracy can be established in the future, it would be possible to improve the accuracy of estimate breeding values for the various selection traits, by using genomic information and phenotypic data in addition to pedigree information [7].
Through a genome-wide association study (GWAS) among populations using linkage disequilibrium (LD), it is possible to detect the relationships between single nucleotide polymorphisms (SNPs) that affect economic traits, while quantitative trait loci (QTL) can be excavated and tagging SNPs selected. Since its first used by [8], polymorphic information content (PIC) had become the most widely applied method for genetic studies to measure the information content of molecular markers. The PIC value of marker is defined as the expected fraction of informative offspring from pedigree [9]. Heterozygosity refers to the ratio of heterozygote in the locus, and observed heterozygosity (OHE) was investigated to analyze the actual heterozygosity degree of each marker for the population. Through the single-step genomic best linear unbiased prediction (ssGBLUP) using an algorithm that combines existing pedigree information with genomic information, expected breeding value (EBV) and genomic expected breeding value (GEBV) can be estimated simultaneously, while through the Back solution, SNPs effects can be estimated [10,11].
For the pigs, studies on the excavation of quantitative traits for economic traits have been performed continually, primarily using GWAS [12–14]. The present experiment was conducted to estimate the SNP effects that affect pH value in the meat of Berkshire pigs.
MATERIALS AND METHODS
A total of 2,037 heads were genotyped using Porcine SNP60k v2 Beadchip (Illumina, San Diego, CA, USA) and 61,565 SNPs were collected. To ensure the quality of the genotypic data, the following SNP types were excluded from the analysis: SNP on sex chromosomes (1,458); SNP without information on chromosomes (7,849); SNP with missing rate higher than 10%; SNP without polymorphism (all homo or hetero); SNP with minor allele frequency lower than 1%; SNP with Hardy-Weinberg disequilibrium chi-squared value greater than 23.93 (p < 10−6); and animals with SNP missing rate higher than 10%. 59 heads were found to have an SNP missing rate higher than 10%. Therefore, the number of animals and SNPs included after quality control was 1,978 and 39,603, respectively (Table 1).
Data on pH values were collected from 882 heads of pigs slaughtered at Namwon Jeil Food between 2015 and 2018. A pH*21K meter (NWK-Binär GmbH., Buchloe, Germany) was inserted into the sirloin muscle and pH values were measured 3 times each at 45 min (pH45m) and at 24 hours (pH24h) after slaughter, and the average values were recorded.
Variance Components: Using the multiple traits animal model, we estimated the variance component and genetic parameters of the pH values, and the equation is as follows:
Where, ytijkl is observed values of t th trait, μ is overall mean, YMti is the ith fixed effect of slaughter year-month, stj is the j th fixed effect of sex, atk is the k th additive genetic effect, etijkl is the residual random effects. We used VCE ver. 6.0 [15] to estimated variance components with pH45m and pH24h.
Estimation of genomic breeding values: The slaughter year-month (YM) and sex were included as fixed effects in a statistical analysis that used the following model:
Where, yi is the vector of observation in i th trait, bi is the vector of fixed effect, ai is the vector of additive genetic random effect, ei is the vector of residual effect, Xi and Zi were known incidence matrix corresponding to bi and ai respectively. Mixed model equation was as follows:
Where, is the inverse matrix of numerator relationship matrix, G−1 is the inverse matrix of genomic relationship matrix, is the inverse matrix of numerator relationship matrix of pigs with genomic information.
SNP effect: The SNP effect of each marker was estimated through reverse operation method of GEBVs and equation was as follows:
Where, is the vector of SNP effect, is the vector of GEBV, Z is the coefficient matrix of SNP, D is the weighted vector [16,17]. We used BLUPF90 family program [18] to estimated SNP effects with pH45m and pH24h.
RESULTS AND DISCUSSION
The average physical distance between adjacent SNP pairs was 61.7 kbp (Fig. 1). The number and proportion of SNPs whose minor allele frequency (MAF) was below 10% were 9,573 and 24.2%, respectively, while those for which it was higher than 40% were 7,343 and 18.5%. The number and proportion of SNPs for which the physical distance between adjacent SNP pairs was less than 1 kbp were 415 and 1.0%, respectively, while those for which it was in the range between 10 kbp and 100 kbp were 30,345 and 76.6%, respectively (Table 2).
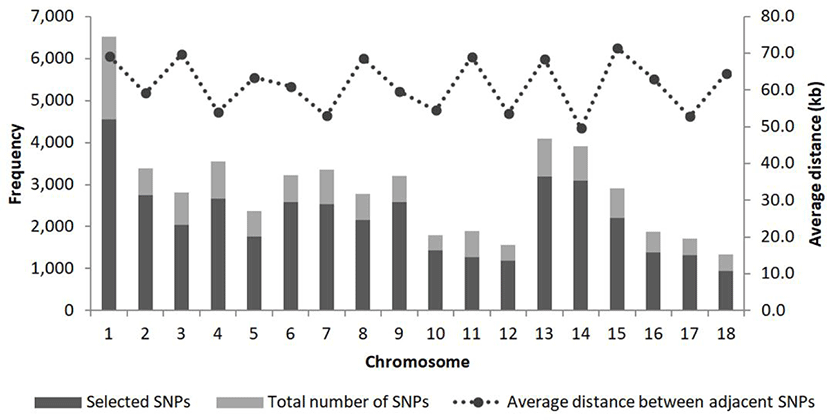
OHE was 0.32 ± 0.16 on average, and in most chromosomes was within the range of 0.31–0.33; it was highest in chromosome 8 (0.36) and lowest in chromosome 15 (0.25). PIC was 0.26 ± 0.11 on average, within the range of 0.22–0.28 (Table 3).
Lee et al. [19] have reported that OHE estimates in Berkshire, Landrace, and Yorkshire were 0.33 ± 0.15, 0.36 ± 0.14 and 0.36 ± 0.14, respectively while estimates for the PIC were on average 0.26 ± 0.11, 0.28 ± 0.10 and 0.29 ± 0.10, respectively. Compared with the results from this experiment with our own, the OHE estimates obtained from our experiment were slightly lower but similar, while PIC estimates were similar.
The estimate for average LD (r2) was 0.40, which was high in the order of chromosomes 1 (0.45), 14 (0.44), 15, and 18 (0.42) (Table 4).
Clearly markers with greater numbers of alleles tend to have higher PIC values and thus are more informative [9]. In the results of this study, it can be seen that the values of MAF, OHE, and PIC tend to be low. This seems to be a result of high immobilization of Berkshire breed used in the study.
Estimates for pH45m and pH24h values were on average 6.40 (± 0.20) and 5.90 (± 0.12), respectively (Table 5). Ryu et al. [20] have reported that in the Berkshire breed the average pH45m estimates in females and males were 6.26 (± 0.08) and 6.15 (± 0.05), respectively, while for pH24h they were 5.61 (± 0.02) and 5.69 (± 0.01) for female and male pigs, respectively. The same study also has reported that pH45m estimates for in females and males were 5.80 (± 0.04) and 5.92 (± 0.05), respectively, in Landrace and 6.05(± 0.02), 5.98 (± 0.03), respectively, in Yorkshire while for pH24h estimates were 5.49 (± 0.01) and 5.51 (± 0.02) in in Landrace females and males and 5.56 (± 0.01), and 5.56 (± 0.01) in Yorkshire pigs. In view of these results, average pH values in the Berkshire breed appear to be higher than in other breeds.
| Traits | Mean ± SD | Min | Max | Skewness |
|---|---|---|---|---|
| pH value after 45 minutes | 6.40 ± 0.20 | 5.94 | 7.09 | 0.22 |
| pH value after 24 hours | 5.90 ± 0.12 | 5.58 | 6.63 | 0.73 |
The genetic parameters of pH45m and pH24h were estimated using VCE6.0 software. Heritability estimates for pH45m and pH24h were 0.10 and 0.15, respectively, while for the phenotypic and genetic correlation between 2 traits they were 0.05 and 0.27, respectively (Table 6). Lee et al. [21] have reported that in the domestic Berkshire breed heritability estimates for pH45m and pH24h were 0.48 and 0.15, respectively. When compare with pH45m and pH24h in this study, the heritability estimates for pH45m obtained from this experiment were lower, while those for the pH24h were similar.
| Trait | h2 ± SE | Correlation1) | |||
|---|---|---|---|---|---|
| 1 | 2 | ||||
| pH45m | 0.0039 | 0.0351 | 0.10 ± 0.04 | - | 0.05 |
| pH24h | 0.0021 | 0.0117 | 0.15 ± 0.07 | 0.27 | - |
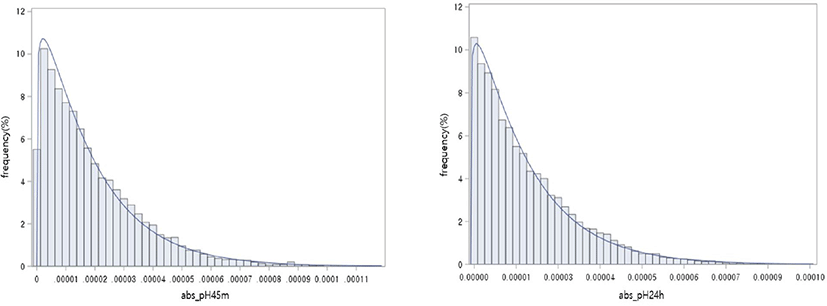
Estimates of SNP effects for pH45m and pH24h were within the ranges of –0.00011536 to 0.00011866 and −0.00009704 to 0.00009562, respectively (Fig. 2). Suitability for normal distribution of SNP effects was tested by 3 methods, but the SNP effects were not normally distributed (Table 7). Absolute values were taken for estimated SNP effects and when expressed as the gamma distribution, most values were near zero, with only a few SNPs having larger effects (Fig. 3).
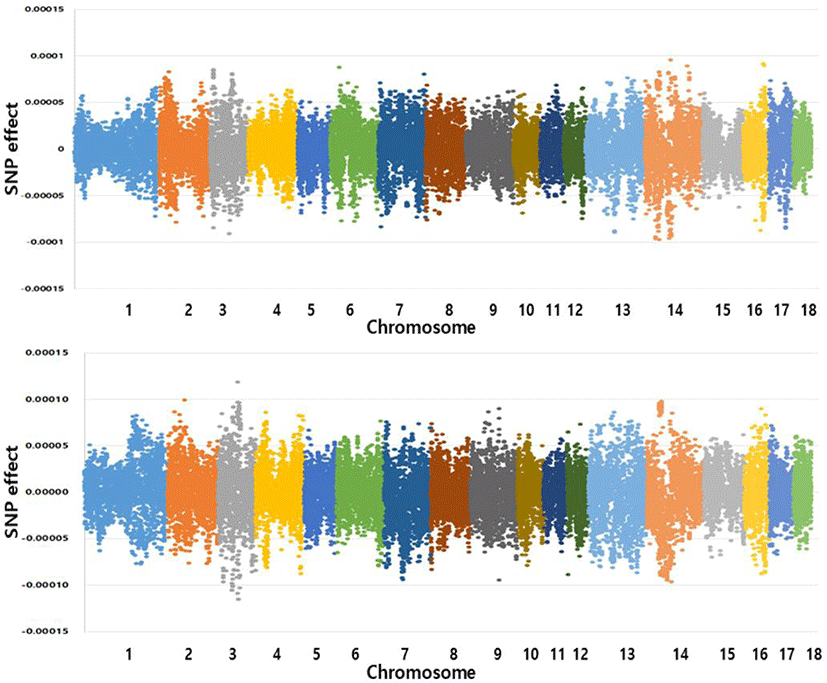
After standardizing estimates for SNP effects, absolute values were taken and were expressed as a Manhattan plot compared to their relative sizes (Fig. 4).
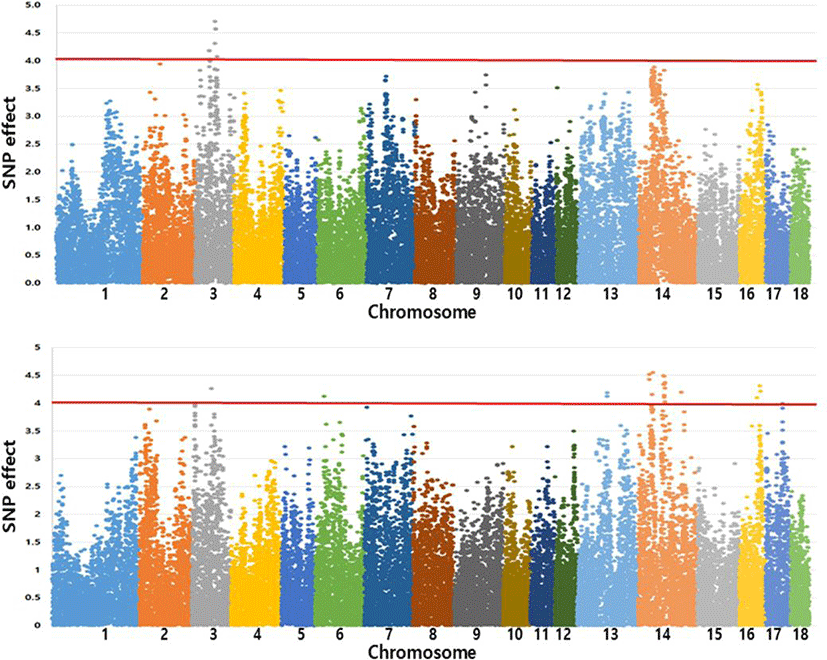
In normal distribution, values within 3 standard deviations of the mean account for approximately 99.74% of the data set. Therefore, SNPs with an absolute value more than 4 standard deviations from the mean were selected as threshold markers. Although the number of SNPs with an absolute value of more than 4 standard deviations was 12 for pH45m and 23 for pH24h (Tables 8 and 9), no significant SNP effects were observed. This may be due to the small number of animals used in the analysis in relation to the number of SNPs.
Six markers (MARC0113402, MARC0100326, MARC0065978, ALGA0115738, ASGA 0015062, and ASGA0015063) that had similar effects nearby markers with significant effects due to LD were detected in chromosome 3 for pH45m, while for pH24h 9 markers were detected (ASGA0063851, MARC0002354, ALGA0078206, ALGA0078209, H3GA0040626, MARC0035949, ALGA0078332, ASGA0064086, and DRGA0013964) in chromosome 14, whose SNPs were in the LD blocks.
Among the SNPs with an absolute value of more than 4 standard deviations, for pH45m and pH24h protein-coding genes were detected in 6 and 4 SNPs, respectively.
For distribution of protein coding genes, a total of 4 genes, <RFX8 (MARC0113402), CREG2 (MARC0100326), TBC1D8 (MARC0065978), and EXOC6B (ALGA0115738, ASGA0015062, ASGA0015063)> were detected in chromosome 3 for pH45m, while for pH24h C12orf49 (ALGA0076917), LOC106506010 (MARC0002354), BICC1 (MARC0035949) and ANK3 (DRGA0013964) were detected in chromosome 14.
Edwards et al. [22] have reported that QTL were discovered for 45-min pH and pH decline on SSC 3 and this QTL region affecting pH on SSC 3 was in a similar location to a pH QTL reported by [23]. When compare with this study, since the markers that we detected in the SSC 3 region are close to or belong to the previously investigated QTL region, it is necessary to study whether they can potentially affect the pH.