
INTRODUCTION
Ewe’s nutrition management during the transition period is a crucial factor influencing lambs’ birth weight, udder development, and milk and colostrum yield. Given that 80% of fetus growth takes place at two last months of pregnancy, the nutrient requirements of ewes increase significantly during this period [1]. Flaxseed (Linumusitatissimum) contains approximately 20% crude protein (CP) and 40% oil on dry matter (DM) basis [2] and it was shown to increase the yield of milk and protein [3]. Feeding animals with flaxseed, as a source of n-3 polyunsaturated fatty acids (PUFA), is also an effective way to improve feed intake and energy balance [4]. Recently, more attention has been paid to flaxseed as a lipid supplement in the ruminant diets due to its high content of α-linolenic acid (over 55% of total fatty acid [FA]) [5], leading to an increase in the concentrations of long-chain fatty acids (LCFA) and PUFA (especially C18:3n3), and decrease the concentration of short-chain fatty acids (SCFA) and medium-chain fatty acids (MCFA) and saturated fatty acids (SFA) in milk fat of dairy cows [6], goats [7] and sheep [8]. Maamouri et al. [9] reported that extruded linseed could block the terminal biohydrogenation steps, thus increasing the ratio of trans-intermediate such as cis-9 trans-11 C18:2 and trans-10 cis-12 C18:2 [10]. Therefore, incorporating flaxseed in ruminant diets can contribute to the prevention of cardiovascular diseases [11] and the modulation of immune and inflammatory responses [12].
It has been reported that high levels of dietary rumen undegradable protein (RUP) and oil can be effective on the dietary protein, milk yield and milk protein efficiencies [3]. Feeding cows with RUP would result in the greater flow of amino acids to the small intestine, increasing intestinal absorption availability [13]. Furthermore, increasing digestion and absorption of proteins in the small intestine following the addition of vegetable oil was previously reported [14]. Therefore, increasing the amounts of dietary RUP and oil or oilseeds may improve the overall use of dietary protein, resulting in increased milk production and protein concentration. Moreover, several studies suggested that providing excess protein in the diet of ewes during late pregnancy is vital for fetus growth, udder development and colostrum and milk yield, and consequently the lamb growth and survival [15,16].
To the best of our knowledge there is little information regarding the interaction between different RUP level and different flaxseed types during transition period of ewes. Thus, we hypothesize that supplementation of ewe’s diets with flaxseeds and RUP in the late gestation may enhance the performance of pregnant ewes. Therefore, the main objective of the present study was to investigate the effects of feeding diets without or with processed flaxseed (whole or extruded) and two theoretical levels of RUP (20% and 40%) on milk yield, milk composition, colostrum yield, milk FAs profile and some plasma metabolites of Baluchi ewes during the transition period.
MATERIALS AND METHODS
This project was carried out at the sheep raising center of Torbat-e-Jam located on the north east part of Iran at 35.2317°N latitude and 60.6401°E longitude from November 2018 to January 2019. All animals were housed and treated following the guidelines suggested by Iranian Council of Animal Care [17]. Seventy-two multiparous Baluchi ewes (48.7 ± 2.8 kg of body weight [BW] and 2–3 years old at the beginning of experiment) were randomly allocated to 6 groups (n =12 ewes in each group) in a completely randomized design with a 3 × 2 factorial arrangements. Before onset the experiment, all ewes were fed flushing diet in August and after breeding they fed on pasture till about 5 weeks prior to lambing. The experiment was conducted from 35-d before parturition (14 d for adaptation to dietary treatments and 21 d for measurements [pre-partum phase]) to day 21 of lactation (post-partum phase). Dietary treatments contained 1) no flaxseed + 20% dietary RUP (no flaxseed, low RUP [NFLR]); 2) no flaxseed + 40% dietary RUP (no flaxseed, high RUP [NFHR]); 3) 10% whole flaxseed + 20% dietary RUP (whole flaxseed, low RUP [WFLR]); 4) 10% whole flaxseed + 40% dietary RUP (whole flaxseed, high RU [WFHR]); 5) 10% extruded flaxseed + 20% dietary RUP (extruded flaxseed, low RUP [EFLR]) and 6) 10% extruded flaxseed + 40% dietary RUP (extruded flaxseed, high RUP [EFHR]). Ewes were housed in the 1.5 × 1.5 m individual tie stalls with rubber mats. The animals had free access to fresh water and feed. Diets were offered as a total mixed ration (TMR) which were formulated based on small ruminant nutrition system (SRNS) [18] to meet the ewe’s nutrient requirements [19]. The diets were fed to ewes at 08:00 and 16:00 h during pre-partum and post-partum periods to ensure about 5% ort.
To achieve isonitrogenous and isoenergetic diets they were formulated by replacing barley grain with flaxseed and nitrogen (the proportions of RUP) balanced using change in the proportions of soybean meal and urea. The Yasminomax product (46% CP and 70% RUP) as a source of RUP was purchased from Sanadam Pars, Tehran, Iran. Ingredients and chemical composition of diets and flaxseed during the transition period are shown in Tables 1 and 2, respectively.
3) Mineral and vitamin mix contained 200 g/kg Ca, 98 g/kg P, 21 g/kg Mg, 44 g/kg Na, 0.3 g/kg Cu, 2 g/kg Mn, 3 g/kg Fe, 3 g/kg Zn, 0.1 g/kg I, 0.1 g/kg Co, 0.001 g/kg Se, 500,000 IU/kg of vitamin A, 100 mg/kg of vitamin E, 100,000 IU/kg of vitamin D3, and 400 mg/kg Antioxidant.
DM, dry matter; RUP, rumen undegradable protein; NFLR, no flaxseed, low RUP; NFHR, no flaxseed, high RUP; WFLR, whole flaxseed, low RUP; WFHR, whole flaxseed, high RU; EFLR, extruded flaxseed, low RUP; EFHR, extruded flaxseed, high RUP; ME, metabolizable energy; CP, crude protein; NDF, neutral detergent fiber; EE, ether extract; RDP, rumen degradable protein.
The DM intake and refusals were recorded daily. All ewes were weekly weighted 4 h before morning feeding. The blood samples were gathered from jugular vein, using heparinized plastic syringes, 3 h after the morning feeding on days 7 and 14 pre- and post-partum to obtain plasma via centrifugation at 3,000×g for 15 min. The plasma was stored at –20°C until later analysis for glucose, β-hydroxybutyric acid (BHBA), non-esterified fatty acid (NEFA) and urea by commercial kits (Pars Azmon, Tehran, Iran and Randox, Randox Laboratories, Crumlin, UK).
To determine colostrum yield, ewes were milked 1, 10 and 18 h post-parturition and a 50-g samples of colostrum were collected and analyzed for fat and protein contents [20]. The ewes were milked twice a day at 09:00 and17:00 h and individual yields were recorded at each morning and evening milking. The collected milk samples from each ewe were mixed proportionally based on morning and evening milk yield and then were analyzed for fat and protein by a MilkoScan (TM minor model 78110, Foss Analytical A/S, Hillerød, Denmark) and also milk FAs profile.
The DM was measured by drying a subsample at 105°C (method no. 934.01) in a forced-air oven [21]. The ash (method no. 945.38) and the ether extract (EE, method no. 945.18) contents were determined based on AOAC procedures [21]. The CP content (method no. 997.06) was measured by kjeldahl method (Kjel-Foss, Kjeltec Auto 1030). The acid detergent fiber (ADF) and neutral detergent fiber (NDF) concentrations were measured according to the methods presented by Van Soest et al. [22].
Plasma metabolites were determined by an auto-analyzer (Abbott Alcyon 300, Abbot Diagnostics, Lake Forest, IL, USA). The profile of milk FAs were determined as described by Fougère et al. [23]. Briefly, the incubation of freeze-dried milk samples was conducted with 2 mL of 0.5 M sodium methoxide in anhydrous methanol and 1 mL of hexane at 50°C for 15 min, and the mixture was cooled. They were then incubated under similar conditions with the addition of 1 mL of 37% methanol/hydrochloric acid (95:5 v/v). To extract fatty acid methyl esters (FAME) of the milk samples, 1.5 mL of hexane and 3 mL of aqueous (6% w/v) potassium carbonate were added and recovered in the hexanoic phase. One µL of recovered hexane phase was injected into gas chromatography (CP-3800 GC, Varian, Palo Alto, CA, USA) assembled with a flame ionization detector at 260°C and a CP-Sil 88 capillary column (100 m × 0.25 mm i.d. × 0.2 μm film thickness; made by Chrompack [Middelburg, The Netherlands], supplied by Varian [Mississauga, ON, Canada]). Helium with a flow rate of 20 cm/sec was used as a carrier gas. The FAME profile was determined in a 1-µL sample at a 1:100 split ratio and 260°C temperature of the injector using a temperature program (140°C [5 min] to 240°C at 4°C/min). To identify peaks, the retention time was compared with commercial standards that contained mixtures of 37 FAME (18919-1AMP, LR-0565, Sigma, St. Louis, MO, USA; and O5632, Sigma, Steinheim, Germany).
All data were analyzed using the MIXED procedure of SAS Institute [24] for a completely randomized design with a 3 × 2 factorial arrangement of treatments. The Duncan’s Multiple Range Test was employed to determine means that were significantly different at p < 0.05. Trends were also considered when p < 0.1. Data were analyzed using the following statistical model:
Where, Yijk are dependent variables, μ is the total mean, Ei is random effect of animal, Fi is fixed effect of flaxseed factor, Rj is fixed effect of RUP factor, (FR)ij is interaction effect of flaxseed and RUP, and eijk is random residual error with a mean and variance of 0 and σ2, respectively.
RESULTS
As shown in Table 3, the dry matter intake (DMI) of ewes was affected by flaxseed, with a significant interaction between flaxseed and RUP level at pre- and post-lambing (p < 0.05). Increasing RUP caused lower DMI in ewes fed a diet without flaxseed but did not affect DMI in flaxseed fed ewes at prepartum. During pre-lambing, ewes fed a diet containing 10% extruded flaxseed showed a higher DMI when compared to other groups (p < 0.01). However, at post-lambing, ewes fed 10% extruded flaxseed (p < 0.05) or 20% RUP (p < 0.01) had significantly higher DMI than those fed no flaxseed or 40% RUP (p < 0.05). At post-lambing, body weight change tended to increase in animals fed the diets containing 40% RUP (p < 0.1) compared to 20% RUP.
Flaxseed had no significant effect on colostrum and its composition, the colostrum yield tended to increase (p < 0.1) in ewes fed the diets containing extruded flaxseed compared to the whole flaxseed or no flaxseed diets (Table 3). Although milk yield was not significantly influenced by flaxseed or RUP, their interaction was significant, and milk yield increased by increasing RUP in ewes fed a diet without flaxseed (p < 0.05), whereas it had no effect on ewes fed with whole or extruded flaxseed. Thus, ewes fed with NFHR, WFHR, EFLR, and EFHR produced higher milk compared to NFLR and WFLR (p < 0.05). The RUP levels influenced the fat and protein concentrations of milk (p < 0.05 and p < 0.01, respectively), and ewes fed low RUP had more fat percentage but produced lower protein compared to those fed high RUP. There was also a significant interaction between flaxseed and RUP for milk fat percentage (p < 0.05).
The serum concentration of NEFA was not affected by the flaxseed or RUP level at pre-lambing (Table 4). However, there was a significant (p < 0.01) interaction between flaxseed and RUP for NEFA concentration at post-lambing. As shown in Table 4, at prepartum, the concentration of BHBA was affected by flaxseed (p = 0.001) and RUP level (p = 0.07) with a significant interaction effect (p = 0.001). In this regard, the serum concentration of BHBA reduced with RUP raising in flaxseed fed ewes; however, a greater BHBA in ewes fed a diet without flaxseed was indicated in the diet with higher RUP level. A converse trend for BHBA was observed postpartum. At the post-lambing period, the BHBA concentrations significantly rose (p < 0.01) in extruded flaxseed fed ewes compared to whole flaxseed or no flaxseed.
Blood glucose and urea concentrations boosted by increasing RUP at pre- and postpartum (p < 0.01). During both lambing periods, the ewes fed extruded flaxseed had a higher glucose concentration than other groups (p < 0.01). The serum concentration of blood urea was also affected by flaxseed type at the pre- and post-lambing periods (p < 0.01), and higher blood urea concentration was observed in animals fed whole flaxseed based diets than other groups.
The effects of dietary treatments on milk FAs composition are presented in Table 5. In general, most FA was affected by flaxseed. The flaxseed supplementation and also RUP level did not significantly affect C4:0 to C12:0 percentages. Higher C13:0 was detected in the ewes fed EFLR when compared to other treatments (p < 0.05). Animals fed the flaxseed diets produced milk containing a lower amount of C14:0, C17:0, C17:1, C20:0, and C21:0 than those on the no flaxseed diets, regardless of RUP level (p < 0.05). However, C14:1, C16:1, and C18:2 (linoleic acid) significantly increased in ewes fed flaxseed (p < 0.05, p < 0.01 and p < 0.01, respectively). The highest concentration of C18:3 (α-linolenic acid) was observed in milk fat of ewes fed extruded flaxseed compared to the other groups, although the ewes fed with whole flaxseed exhibited higher C18:3 than control group (p < 0.05) (Fig. 1). With the exception of C13:0, C17:0, and C22:0, other milk FA were not affected by RUP level. Furthermore, a significant interaction between flaxseed and RUP level was observed for C13:0 and C22:0 (p < 0.01).
short-chain fatty acids (C4:0 to C13:0, SCFA); medium-chain fatty acids (C14:0 to C16:1, MCFA); long-chain fatty acids (> C17, LCFA); polyunsaturated fatty acids (trans-9 trans-12 C18:2, C18:2 n6, C18:3n3, C20:4n6, PUFA); monounsaturated fatty acids (C14:1, C16:1, C17:1,C18:1n9t, cis-9 C18:1, C21:0, MUFA); saturated fatty acids (C4:0, C6:0, C8:0, C10:0, C11:0, C12:0, C13:0, C14:0, C15:0, C16:0, C17:0, C18:0, C20:0, C21:0, C22:0, SFA).
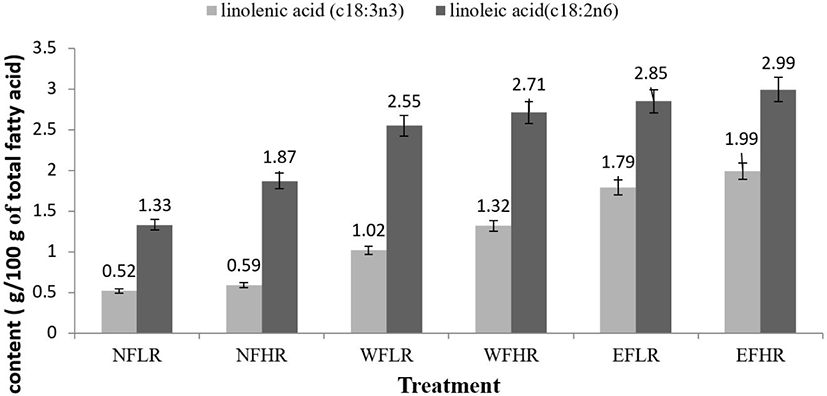
Short and long-chain FA and also monounsaturated FA were not influenced by flaxseed or RUP level, whereas a trend for a lower amount of medium-chain FA was found in extruded flaxseed fed animals compared to those fed a diet without flaxseed (p = 0.09). Feeding ewes with both types of flaxseed significantly increased PUFA concentrations of milk (p < 0.01), and this considerably indicated in ewes fed extruded flaxseed. Taken as a whole, saturated FA, which makes up the principal group, was significantly affected by flaxseed (p < 0.05), and the ewes fed extruded flaxseed indicated a lower concentration of SFA compared to whole, or no flaxseed fed ewes.
DISCUSSION
During pre- and post-lambing, the ewes fed the diet containing 10% extruded flaxseed showed higher DMI than those fed the control diet. In agreement with our results, the inclusion of low to moderate levels of flaxseed, either as whole or extruded (up to 10% of diet DM), increased [6, 25 (at postpartum)] or did not affect DMI [7,25 (at prepartum)] in dairy cows and goats, whereas high inclusion levels (21% of diet DM) decreased DMI of cows [26]. This discrepancy among different studies could be due to different amounts and types of supplemented oilseeds and could be due to their palatability [25]. In general, it was reported that high FA intake could directly suppress DMI due to its inhibitory effect on rumen motility [27] when total fat concentration is higher than 6% of the DM [28].
Reducing DMI by rising the RUP level in the postpartum period in the present study was consistent to the findings of Rehman et al. [13], who reported that the DMI of cows significantly decreased by increasing of RUP level from 30% to 60%. This reduction could be attributed to the presence of less fermentable protein and the subsequent decline in the ruminal ammonia nitrogen, which would reduce the growth rate of ruminal microorganisms, thus diminishing the nutrient digestibility and DMI [29]. Similarly, Hartwell et al. [30] observed that prepartum DMI was not affected by RUP level, while postpartum intake decreased in high RUP diet compared to low RUP diet. However, they indicated that negative effects for postpartum intake following feeding excess protein during late gestation were not a result of rumen degradable protein (RDP) deficiency as they provided similar RDP (but different CP content) for all treatments.
A tendency for greater colostrum production (p < 0.1) in extruded flaxseed fed ewes might be related to higher DMI. Consistent to our results, the inclusion of fish oil as a source of n-3 PUFA did not affect the main constituent of goat’s colostrum [31]. In contrast, increasing the feeding level of fish oil up to 40 g/d led to a linear decline in ewes’ total colostrum output [32]. Supplementation of the ewes’ diets with digestible undegradable protein (DUP) in the late pregnancy resulted in a higher colostrum yield and yields of components (protein, fat, and solid non-fat contents) within 24 h of lambing [15]. However, in another study, Annett et al. [20] found that the negative effect of fish oil on colostrum secretion was alleviated by supplying additional DUP, suggesting the responses to elevated DUP is dependent on metabolizable energy (ME) intake.
In no flaxseed and whole flaxseed diets, increasing RUP level led to numerically increase (p > 0.05) in the milk production by 41.6% and 27.7%, respectively, but extruded flaxseed diets lead to a significant interaction between flaxseed and RUP level which was higher in ewes fed on EFLR compared to WFLR and NFLR. Do Perdo et al. [33] reported that despite the positive effects of feeding flaxseed or linola (4.8%) on DMI and energy balance, milk yield was higher for cows fed Megalc (1.1% of diet DM) which disagrees with the results of Petit [34], who found no difference in milk yield when cows were fed with whole flaxseed (13.9% of diet DM). Conversely, Zachut et al. [24] observed higher milk yield in cows fed 10.7% extruded flaxseed compared to control. Therefore, responses to flaxseed supplementation are still controversial and can be associated to the amounts and forms of supplemental flaxseed or to interactions with other diet components [25].
In contrast to our findings, supplementation of dairy cow’s diet with whole or micronized flaxseeds reduced milk protein [35] or fat contents [36]. Morsy et al. [37] reported that a decrease in milk fat content following the inclusion of flaxseed oil was related to changes in microbial activity and biohydrogenation of PUFA, leading to the accumulation of trans-10 C18:1. The latter has a crucial inhibitory role in short- and medium-chain FA synthesis in the udder’s epithelial cells, hence reducing milk fat [36]. Furthermore, oilseeds are typically rich in long-chain unsaturated FA, which can depress milk fat content through their adverse impacts on fiber digestion and subsequently ruminal acetate concentration [38].
Compared to 20% RUP, ewes consuming 40% RUP produced more milk protein concentration. It was reported that supplementation of ruminant diets with RUP could enhance the flow of nitrogen and essential amino acids to duodenum; hence, increase the milk yield and milk protein concentration in cattle and sheep [13,39]. However, Mikolayunas-Sandrock et al. [40] reported a 14% increase in milk yield and milk fat as well as a 15% increase in milk protein yield when dairy ewes were fed the diets containing high RUP compared to the group receiving lower RUP, without any significant changes in milk fat or protein percentages.
Almost all measured blood metabolites were within the range reported for sheep [41,42]. Except for NEFA at prepartum, other blood parameters were significantly affected by flaxseed, RUP, or their interactions. Petit [27] found no changes in NEFA, BHBA, and glucose concentrations for cows fed with different levels of whole flaxseed. He noted that this lack of effect was related to a similar energy balance among treatments. The higher concentration of BHBA in ewes fed NFHR could be ascribed by lower prepartum energy intake which could lead to moderate ketosis (0.8 and 1.6 mmol/L), although their blood glucose concentration was within the normal range of 31 to 81 mg/dL reported by Christian and Pugh [43]. It has been stated that, in contrast to dairy cows, an increased concentration of ketone bodies is not always accompanied by glucose deficiencies in ewes [44]. However, at post lambing, concentrations of BHBA in NFLR fed ewes were back to normal range. An increase in serum concentration of glucose and the significant reduction of serum BHBA in ewes fed WFHR and EFHR at the pre-lambing period indicated the improved energy status of body compared to other treatments.
The serum NEFA concentration is considered an index for adipose fat mobilization [45]. Increased NEFA concentrations postpartum in ewes fed whole or extruded flaxseed compared to the control might indicate the breakdown of fat as a result of increased energy demand, suggesting that the ewes were in negative energy balance due to the increased nutrient demands for milk production. However, NEFA concentration may progressively fall due to increased DMI and energy balance [46]. Bertics et al. [47] reported that DMI is inversely related to concentrations of NEFA and BHBA in plasma which disagrees with the data from the current study. Kholif et al. [48] reported that plasma concentrations of NEFA were increased with feeding 20 mL flaxseed or soybean oil; however, in another study, the inclusion of 50 g crushed flaxseed or 20 mL flaxseed oil did not affect NEFA concentration in goats [7]. This discrepancy may be due to providing additional energy density required for milk production.
The diets containing extruded flaxseed and also 40% RUP elevated the serum glucose concentration in ewes at pre- and post-lambing periods which is in agreement with Jahani-Moghadam et al. [49] and Kholif et al. [48], who reported higher serum glucose concentration by adding flaxseed or feeding diets containing n3-PUFA. This increment might be due to the improvement in nutrient digestion and ruminal fermentation, as well as improved production of propionate [37,48]. Furthermore, it seems that blood glucose concentration is related to changes in DMI as well as the effect of supplemented extruded flaxseed as a source of PUFA on glucose metabolism. Qin et al. [50] reported that the increase in glucose concentrations in fat supplemented groups might be as a consequence of higher somatotropin concentrations, as somatotropin was found to stimulate hepatic gluconeogenesis to supply the energy demand of the lactating mammary gland [51]. Moreover, increased RUP resulted in higher serum glucose content through gluconeogenesis from excess amino acids other than its utilization towards mammary protein synthesis. Therefore, it may explain the increase of serum concentration of glucose caused by increasing RUP level to 40% in treatments containing extruded flaxseed. These findings are in agreement with those presented by Milis et al. [52] and Amanlou et al. [15].
Plasma urea is an indicator of protein supply or protein utilization that is influenced by the animals’ nutritional status [53]. Reduction in blood urea concentration in ewes fed extruded flaxseed compared to those fed with whole flaxseed or without flaxseed may probably be explained by the effectiveness of oilseeds in controlling the ruminal protozoa population and improving the efficiency of dietary protein intake [54]. Furthermore, Johnson et al. [55] stated that feeding cows with oilseeds increased plasma urea levels due to increased ruminal nitrogen absorption. The significant increase in serum blood urea concentration with a higher RUP level of diets to 40% was somewhat unexpected. However, this finding is in agreement with Amanlou et al. [15], who reported that elevating DUP and dietary protein simultaneously resulted in increased serum blood urea of sheep.
It has been well documented that diet strongly affects both milk fat content and milk FA profile [56]. Similar to our results, Zachut et al. [24] and Neveu et al. [6] observed that the inclusion of flaxseed did not affect the short-chain FA (C4 to C10) content of cow’s milk fat. In another study, Glasser et al. [57] reported that oilseeds rich in C18:3 like flaxseed had lower inhibitory effects on de novo short-chain FA synthesis compared to oilseeds rich in C18:2. The abundance of unsaturated FAs, especially C18:3, would explain the results obtained in whole or extruded flaxseed diets. The intermediates of these unsaturated FA which are produced during biohydrogenation in the rumen, can inhibit de novo FA synthesis in the mammary gland and would cause a relative reduction in short and medium-chain FA [56], as seen in Table 5. In the current study, a decrease in odd and branched-chain FA (C15:0 and C17:0) following the inclusion of extruded or whole flaxseed is consistent to Isenberg et al. [58]. This reduction could be due to the variation in the rumen bacterial population since most of these FA derived from bacteria leaving the rumen [59].
The higher concentrations of C14:1 and C16:1 in milk fat along with higher Δ9-desaturation indexes of C14 and C16 (data not shown) in ewes fed extruded and/or whole flaxseed diets than the control group suggested an increase in Δ9-desaturase activity. However, Correddu et al. [60] reported that despite increasing the concentrations of C14:1, C16:1, and C18:1 with the inclusion of grape seed and linseed in the diet of dairy sheep, the desaturase indexes did not follow the same trend, indicating the increase of these unsaturated FA were not related to an increase in Δ9-desaturase activity.
An increase in ewes’ milk α-linolenic acid in the diets containing flaxseed agreed with previous studies that used whole or ground flaxseed [5], extruded flaxseed [6], and crushed flaxseed [7]. These elevated effects could simply be a consequence of the direct incorporation of these FA from diet to the mammary gland. Similar to our results, Maamouri et al. [9] reported an increase of 21% of total FA of α-linolenic acid in the diets supplemented with extruded linseed compared to the whole one. Kennelly [61] and Neveu et al. [6] reported that extrusion might denature the protein matrix around the fat droplet, hence protect FAs from ruminal biohydrogenation. However, more recently, Maamouri et al. [9] stated that extrusion might lead to the partial release of the oil due to reduction in the protective effect of oilseeds proteins. This free oil then could interfere with microorganisms responsible for biohydrogenation, thus increase trans-intermediates. Although we could not measure all trans-FAs due to technical limitation, similar concentrations of trans-9 C18:1 and trans-9 trans-12 C18:2 and also more PUFA in milk fat of extruded compared to whole flaxseed fed ewes might indicate the lack of effect of oil on ruminal biohydrogenation.
Decreases in saturated FA in ewes fed extruded flaxseed are in agreement with most experiments on oil and/or oilseeds in ewes [55,62] and cows [6, 24]. Kholif et al. [7] reported that lower total SFA content in animals fed with a diet containing PUFA-rich oil sources was justified by the inhibitory effect of PUFA on de novo FAs synthesis. This increment in PUFA, along with decreased in SFA of milk fat are beneficial in human health in terms of lowering total and low-density lipoprotein cholesterol [63].
CONCLUSION
The results indicated that the supplementation of ewe’s diets with flaxseed (especially the extruded form) during the transition period increased DMI and colostrums compared to diets without flaxseed. However, the advantage of increasing RUP level was to improved milk protein percentage along with higher blood glucose concentration, although ewes fed low RUP had normal glucose contents. Blood parameters varied extensively among treatments due to significant interaction between flaxseed and RUP. The concentrations of linoleic, linolenic, and PUFA were significantly increased about twofold by the inclusion of flaxseed in the diet. In general, it can be concluded that using 10% extruded flaxseed compared to a control diet in transition dairy sheep feeding had beneficial effects on colostrum yield and insignificant but considerable amount of milk production (1.66 vs. 2.03) and also favorable modification of milk FA profile. However, due to similar performance responses to RUP level and economic point of view, it is recommended to use 20% RUP in both pre- and post-lambing.